

An in silico down-scaling approach uncovers novel constituents of the Plasmodium-containing vacuole
- PMID: 30232409
- PMCID: PMC6145888
- DOI: 10.1038/s41598-018-32471-6
An in silico down-scaling approach uncovers novel constituents of the Plasmodium-containing vacuole
Abstract
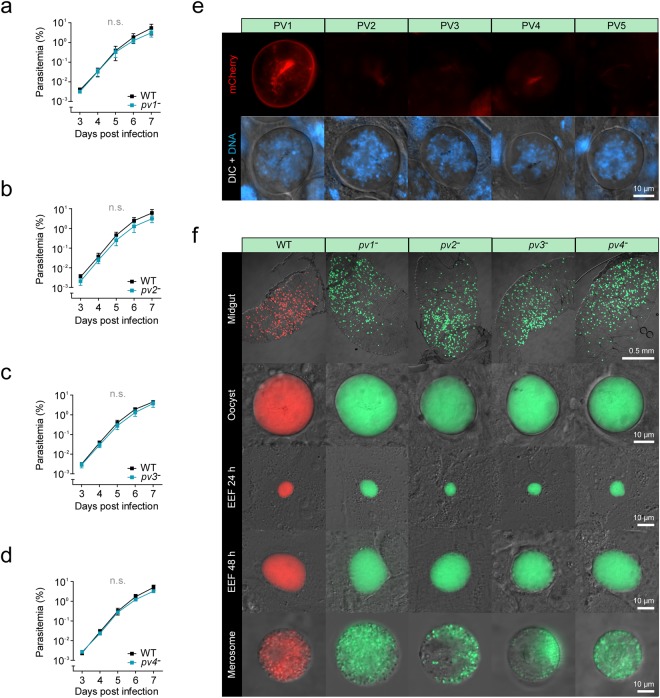
During blood stage development the malaria parasite resides in a membrane-bound compartment, termed the parasitophorous vacuole (PV). The reasons for this intravacuolar life style and the molecular functions of this parasite-specific compartment remain poorly defined, which is mainly due to our limited knowledge about the molecular make-up of this unique niche. We used an in silico down-scaling approach to select for Plasmodium-specific candidates that harbour signatures of PV residency. Live co-localisation of five endogenously tagged proteins confirmed expression in the PV of Plasmodium berghei blood and liver stages. ER retention was ruled out by addition of the respective carboxyterminal tetrapeptides to a secreted reporter protein. Although all five PV proteins are highly expressed, four proved to be dispensable for parasite development in the mammalian and mosquito host, as revealed by targeted gene deletion. In good agreement with their redundant roles, the knockout parasites displayed no detectable deficiencies in protein export, sequestration, or PV morphology. Together, our approach improved the catalogue of the Plasmodium PV proteome and provides experimental genetics evidence for functional redundancy of several PV proteins.
Conflict of interest statement
The authors declare no competing interests.
Figures
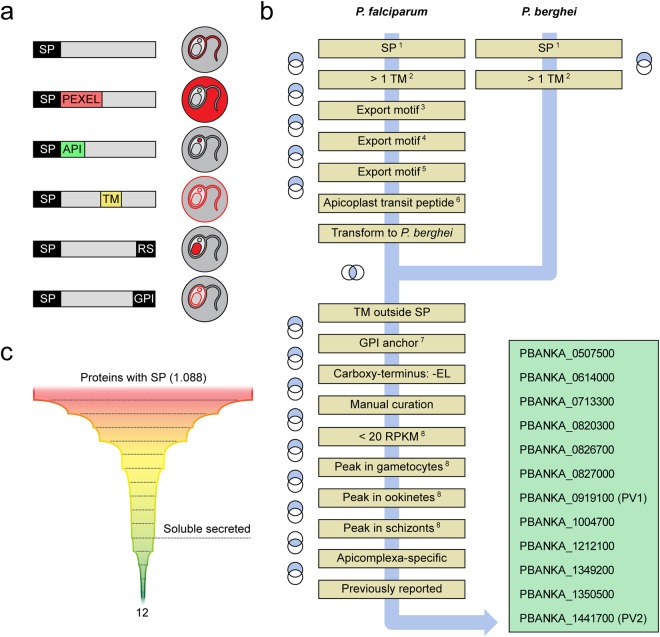
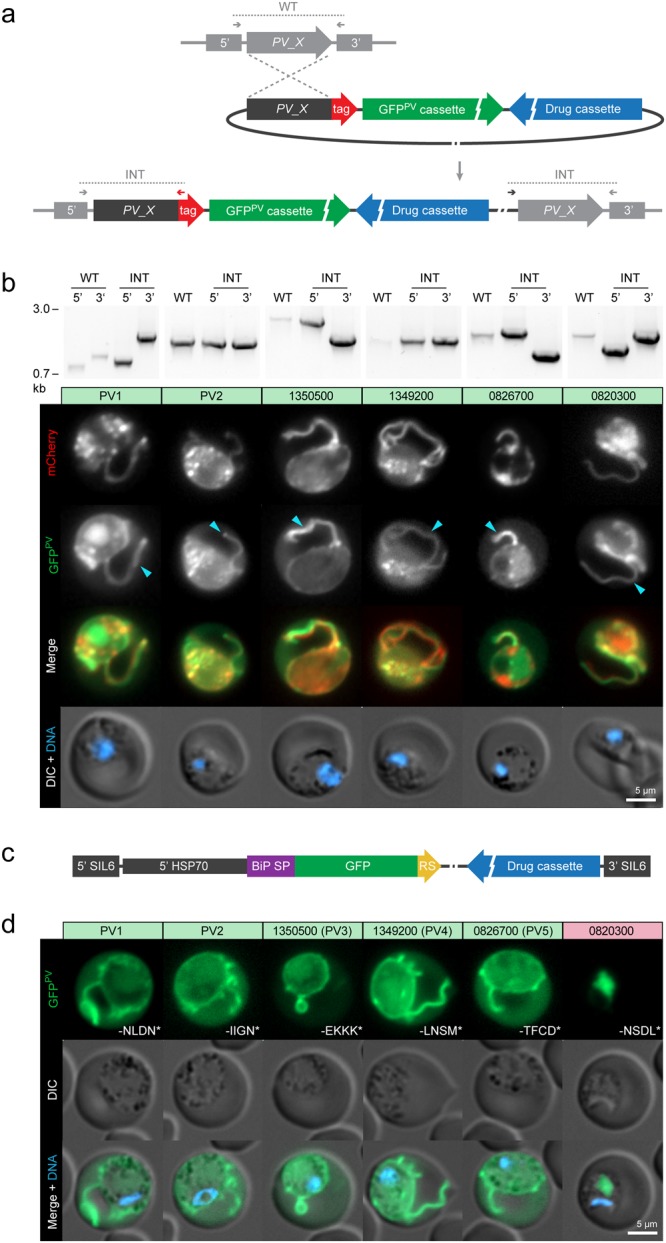
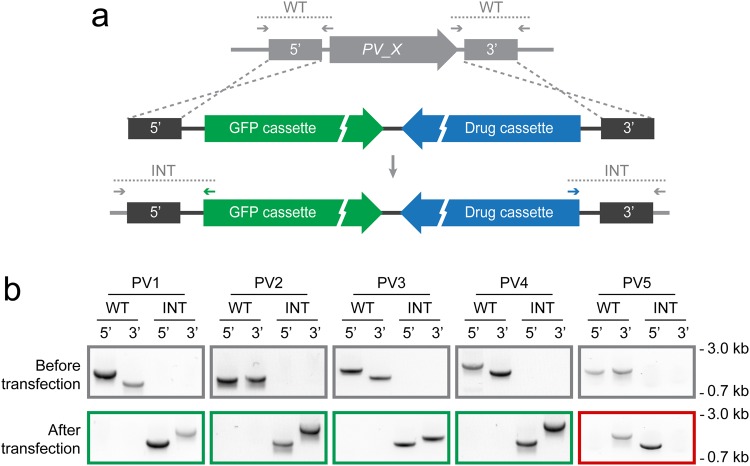
 , PV protrusions. Candidates showing no PV localisation are depicted in Supplementary Fig. S1. Wild-type (WT) and integration-specific (INT) diagnostic PCRs of the parental transgenic parasite lines, as indicated in a and Supplementary Fig. S1, are shown above and indicate proper genomic integration of the targeting constructs. For full size images of DNA gels, see Supplementary Fig. S5. (c,d) Assessment of ER retention. (c) Schematic representation of the vectors used for ER retention testing. The GFPPV cassette, consisting of the signal peptide of BiP (purple) fused to GFP (green), was appended with the last four amino acids of the PV candidates (yellow) and expressed from the silent intergenic locus on chromosome 6 (SIL6) under the control of the HSP70 promoter. (d) Live fluorescence microscopy of transgenic P. berghei parasites expressing ER retention testing constructs. Shown are the GFP signal (green, top), differential interference contrast images (DIC, middle), and a merge with Hoechst 33342 nuclear dye (DNA, blue, bottom). The white labelling indicates the last four amino acids of the PV candidates.
, PV protrusions. Candidates showing no PV localisation are depicted in Supplementary Fig. S1. Wild-type (WT) and integration-specific (INT) diagnostic PCRs of the parental transgenic parasite lines, as indicated in a and Supplementary Fig. S1, are shown above and indicate proper genomic integration of the targeting constructs. For full size images of DNA gels, see Supplementary Fig. S5. (c,d) Assessment of ER retention. (c) Schematic representation of the vectors used for ER retention testing. The GFPPV cassette, consisting of the signal peptide of BiP (purple) fused to GFP (green), was appended with the last four amino acids of the PV candidates (yellow) and expressed from the silent intergenic locus on chromosome 6 (SIL6) under the control of the HSP70 promoter. (d) Live fluorescence microscopy of transgenic P. berghei parasites expressing ER retention testing constructs. Shown are the GFP signal (green, top), differential interference contrast images (DIC, middle), and a merge with Hoechst 33342 nuclear dye (DNA, blue, bottom). The white labelling indicates the last four amino acids of the PV candidates.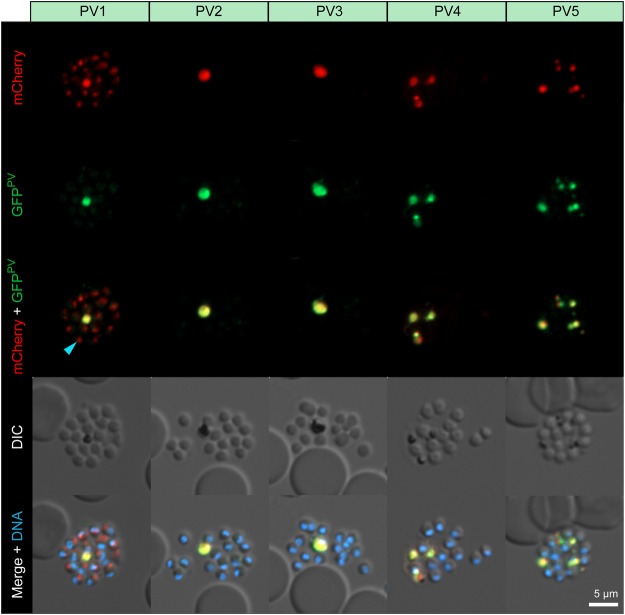
 , PV1-mCherry-fluorescent dot associated with an emerging merozoite.
, PV1-mCherry-fluorescent dot associated with an emerging merozoite.

References
-
- Lingelbach K, Joiner KA. The parasitophorous vacuole membrane surrounding Plasmodium and Toxoplasma: an unusual compartment in infected cells. J. Cell Sci. 1998;111:1467–1475. - PubMed
-
- Ward GE, Miller LH, Dvorak JA. The origin of parasitophorous vacuole membrane lipids in malaria-infected erythrocytes. J. Cell Sci. 1993;106:237–248. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- intramural funding/Humboldt-Universität zu Berlin (Humboldt-Universität)/International
- intramural funding/Humboldt-Universität zu Berlin (Humboldt-Universität)/International
- GRK2290/Deutsche Forschungsgemeinschaft (German Research Foundation)/International
- GRK2290/Deutsche Forschungsgemeinschaft (German Research Foundation)/International
- intramural funding/Max-Planck-Gesellschaft (Max Planck Society)/International
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
