

An Insect Prostaglandin E2 Synthase Acts in Immunity and Reproduction
- PMID: 30233407
- PMCID: PMC6131586
- DOI: 10.3389/fphys.2018.01231
An Insect Prostaglandin E2 Synthase Acts in Immunity and Reproduction
Abstract
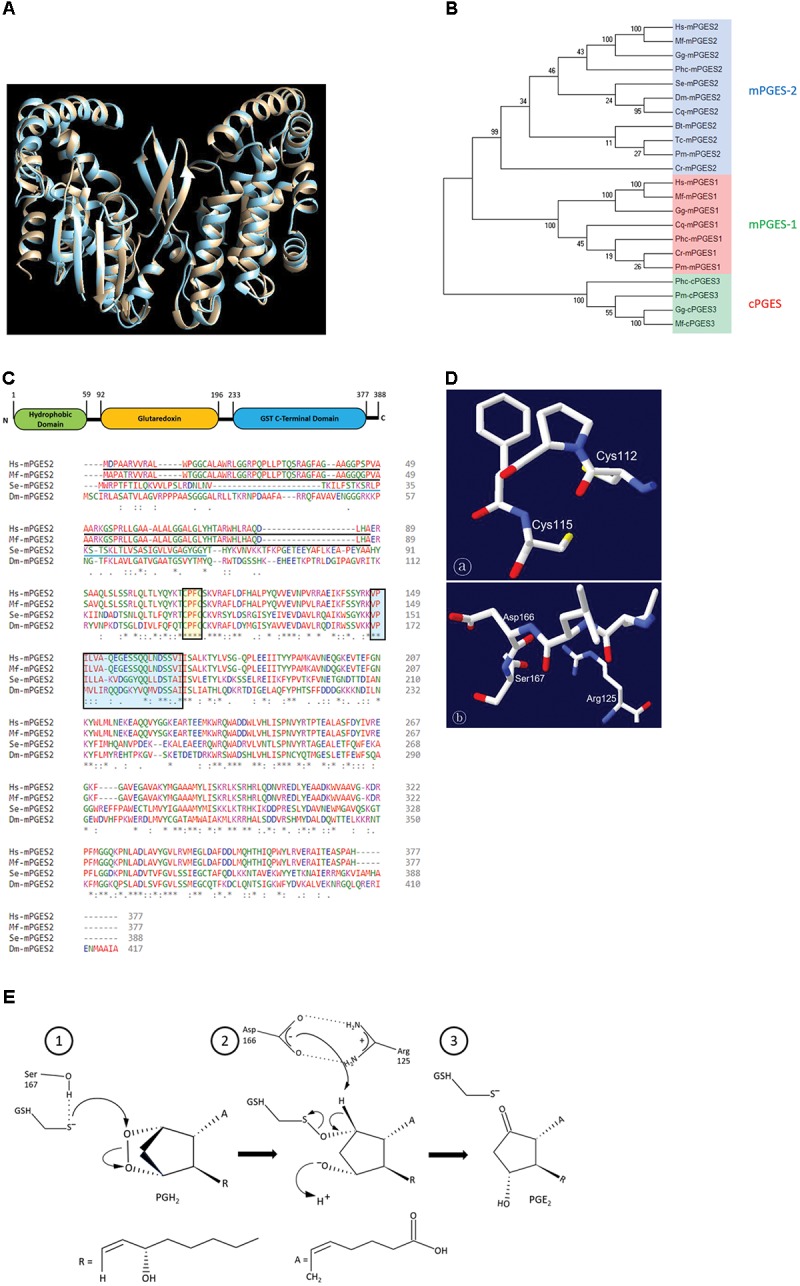
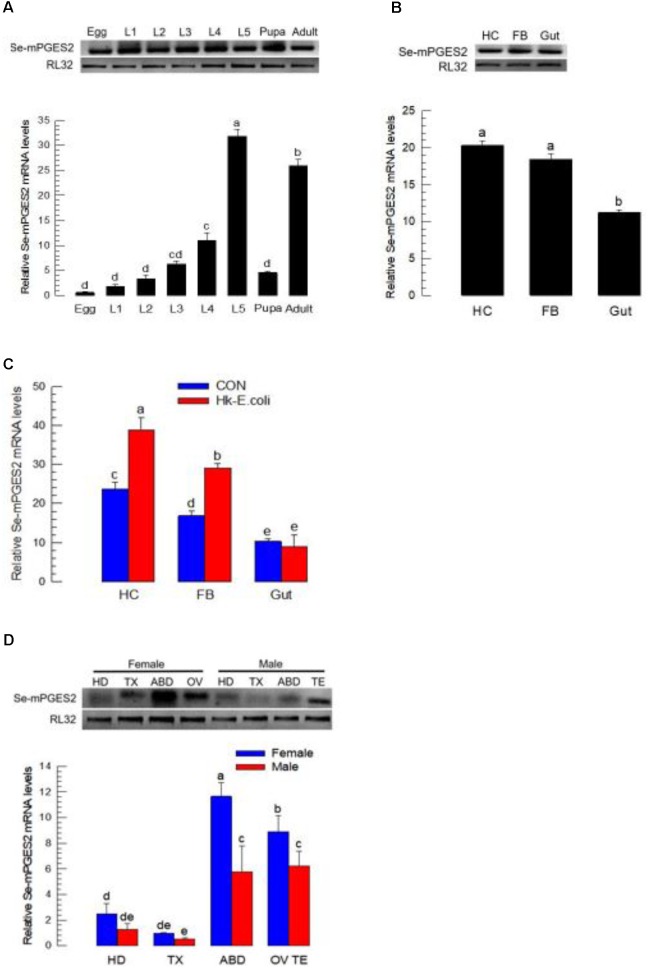
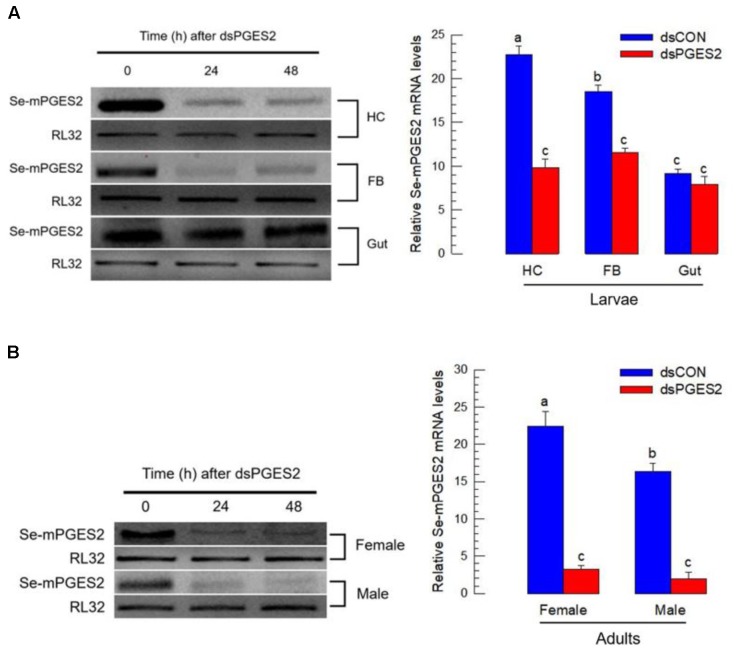
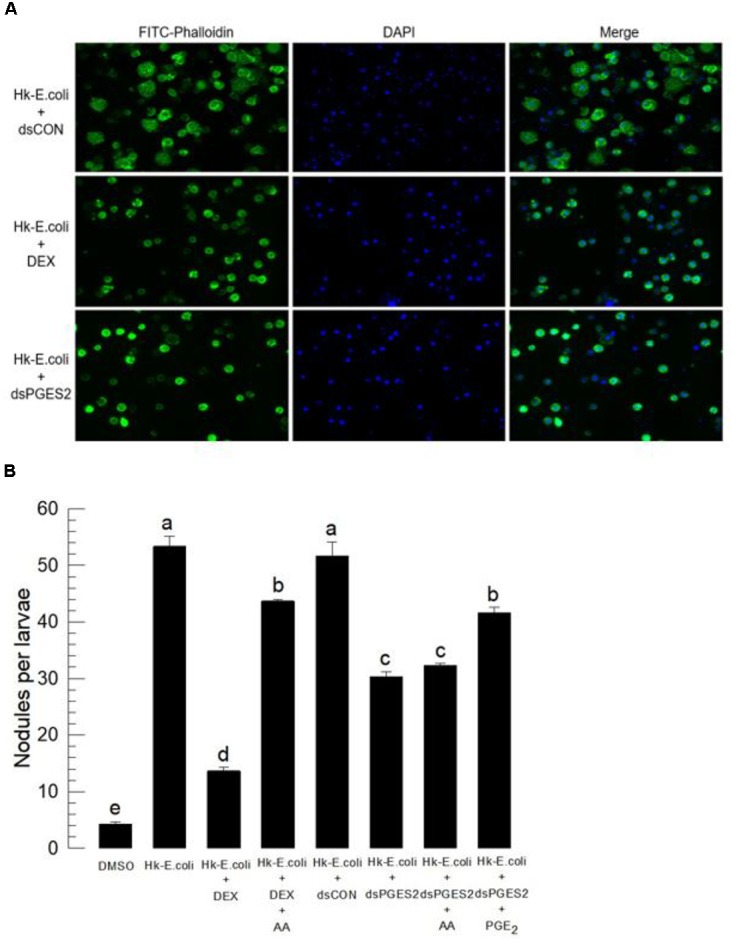
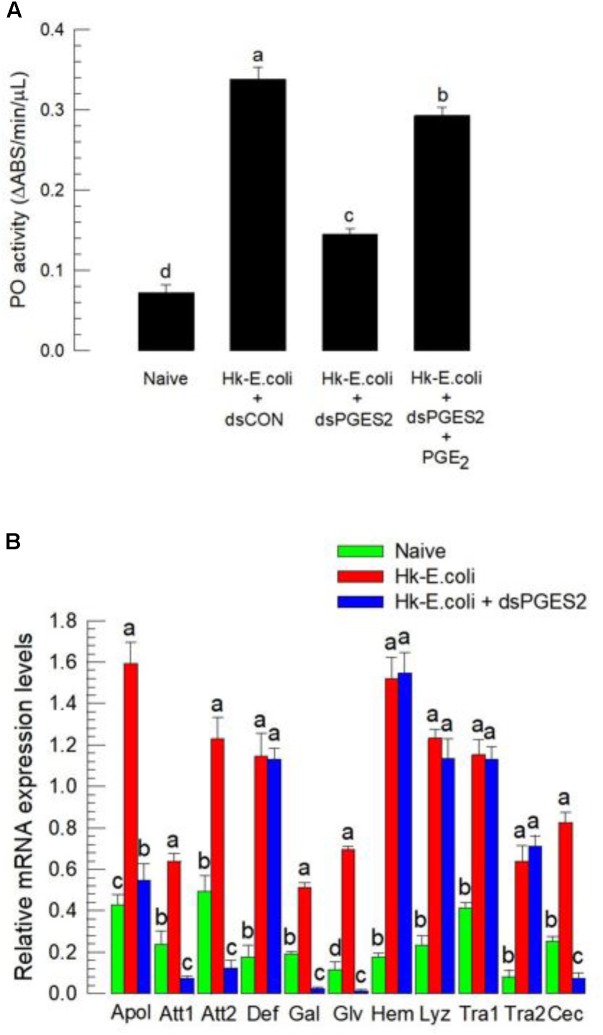
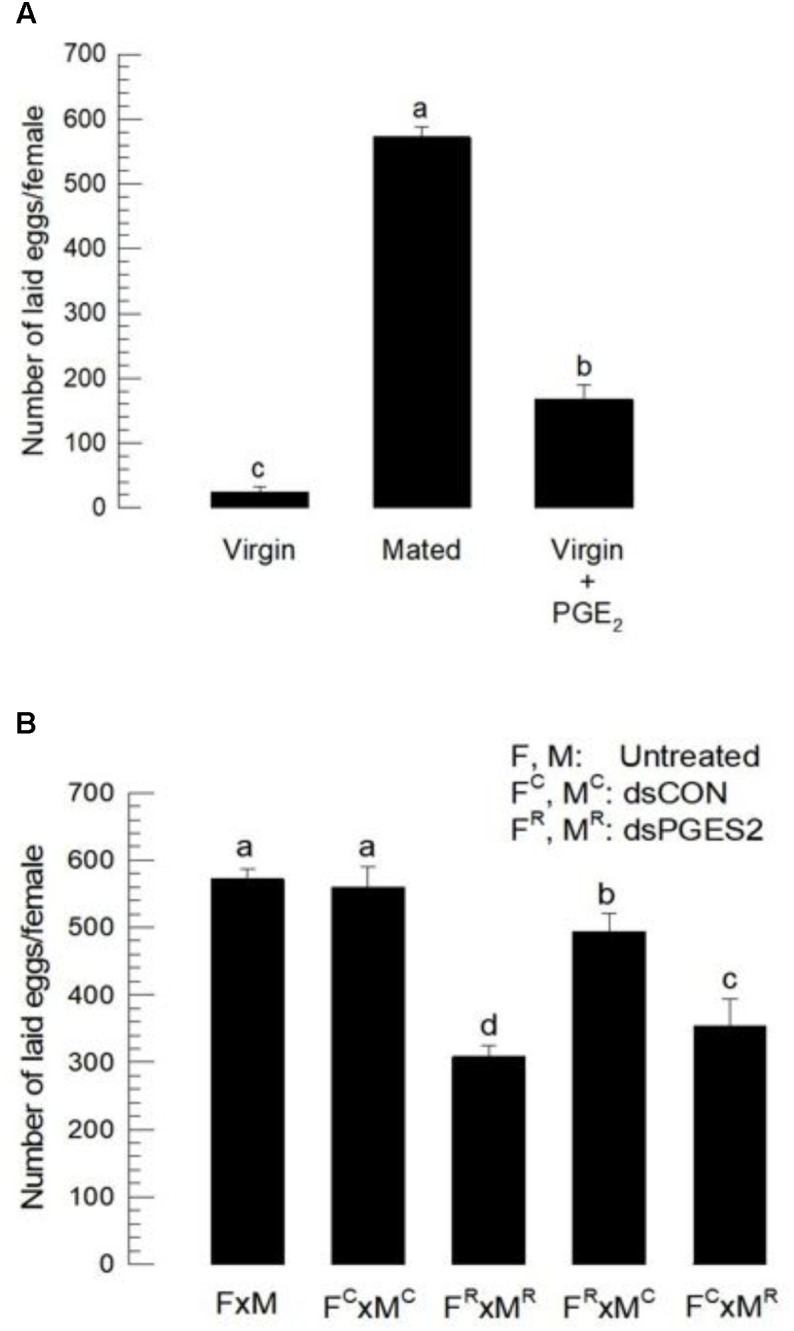
Eicosanoids, oxygenated metabolites of C20 polyunsaturated fatty acids (PUFAs), mediate fundamental physiological processes, including immune reactions and reproduction, in insects. Prostaglandins (PGs) make up one group of eicosanoids, of which PGE2 is a relatively well-known mediator in various insect taxa. While PG biosynthesis has been reported, the specific biosynthetic pathway for PGE2 is not known in insects. Here, we posed the hypothesis that Se-mPGES2 mediates biosynthesis of physiologically active PGE2 through its cognate protein. To test this hypothesis, we interrogated a transcriptome of the lepidopteran insect, Spodoptera exigua, to identify a candidate PGE2 synthase (Se-mPGES2) and analyzed its sequence and expression. Its predicted amino acid sequence contains a consensus thioredoxin homology sequence (Cys-x-x-Cys) responsible for catalytic activity along with an N-terminal membrane-associated hydrophobic domain and C-terminal cytosolic domain. It also shares sequence homology (36.5%) and shares almost overlapping three dimensional structures with a membrane-bound human PGES2 (mPGES2). Se-mPGES2 was expressed in all developmental stages with high peaks during the late larval instar and adult stages. Immune challenge significantly up-regulated its expression levels in hemocytes and fat body. Injecting double-stranded RNA (dsRNA) specific to Se-mPGES2 significantly impaired two cellular immune responses, hemocyte-spreading behavior and nodule formation following bacterial challenge. Humoral immunity was also significantly suppressed, registered as reduced phenoloxidase activity and antimicrobial peptide expression levels. The suppressed immune responses were reversed following PGE2, but not arachidonic acid (AA), treatments. RNAi treatments also reduced the egg-laying behavior of females. Control females mated with the RNAi-treated males led to substantially reduced egg-laying behavior, which was also reversed following PGE2 injections into females. These results strongly bolster our hypothesis that Se-mPGES2 acts in the biosynthesis of PGE2, a crucial biochemical signal mediating immune and reproductive physiology of S. exigua.
Keywords: PGE2; PGES; Spodoptera exigua; eicosanoids; immunity; reproduction.
Figures
References
-
- Bichon A., Boukhatem N., Gay P., Dru P., Terzian H., Petitjean A. M., et al. (2001). Genetic and molecular features of Su(P), a gene that interacts with ref(2)P in male fertility of Drosophila melanogaster. Mol. Genet. Genomics 265 354–361. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources