

Kappa Opioid Receptor Distribution and Function in Primary Afferents
- PMID: 30236284
- PMCID: PMC6300132
- DOI: 10.1016/j.neuron.2018.08.044
Kappa Opioid Receptor Distribution and Function in Primary Afferents
Abstract
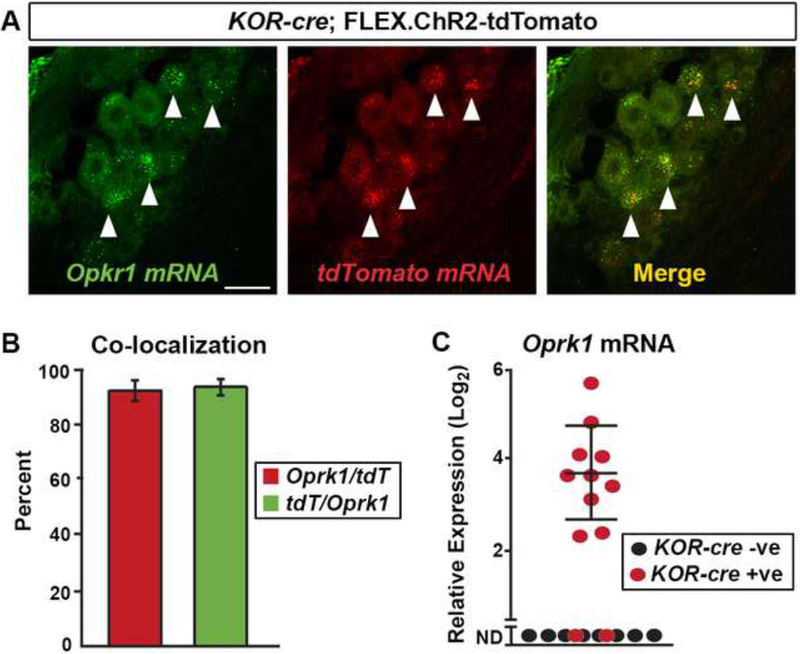
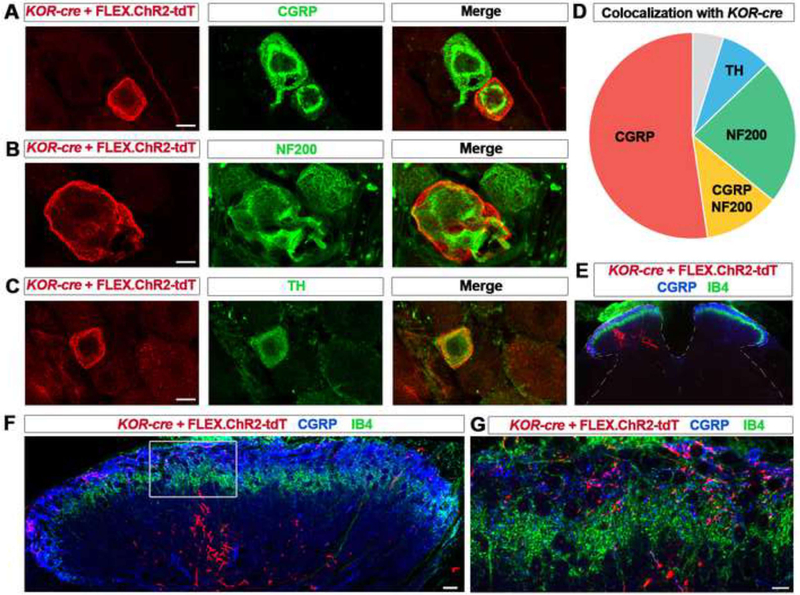
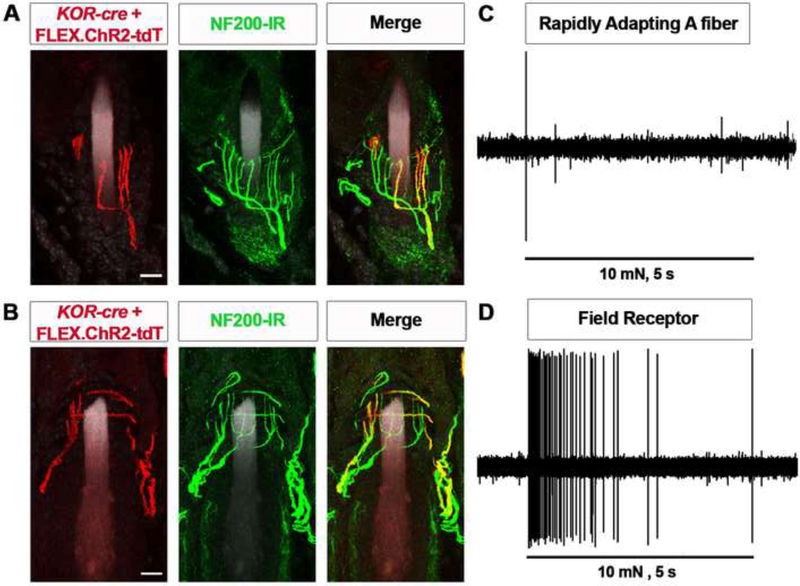
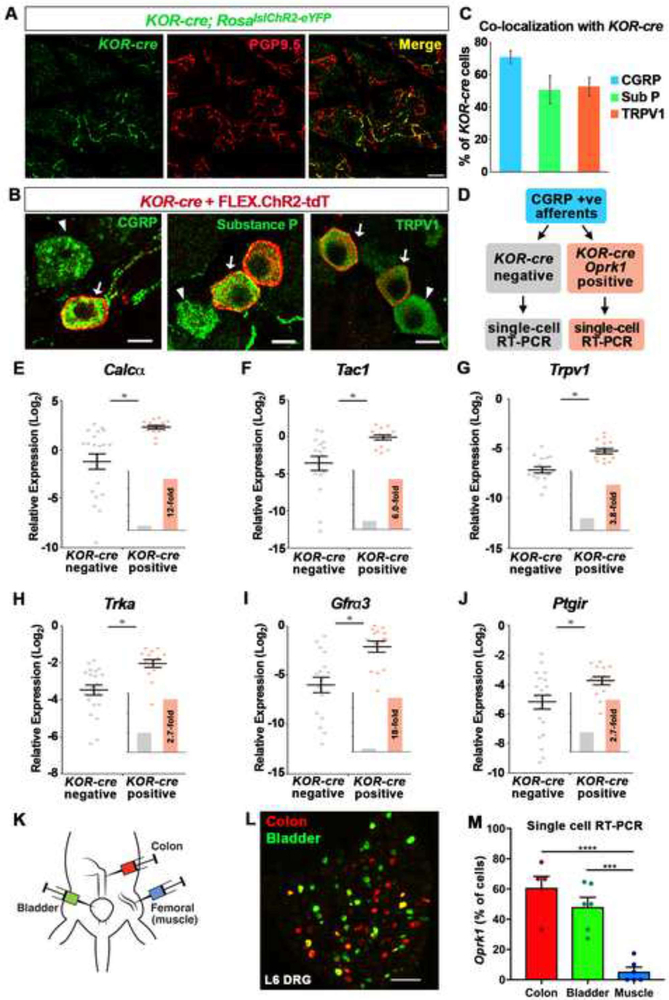
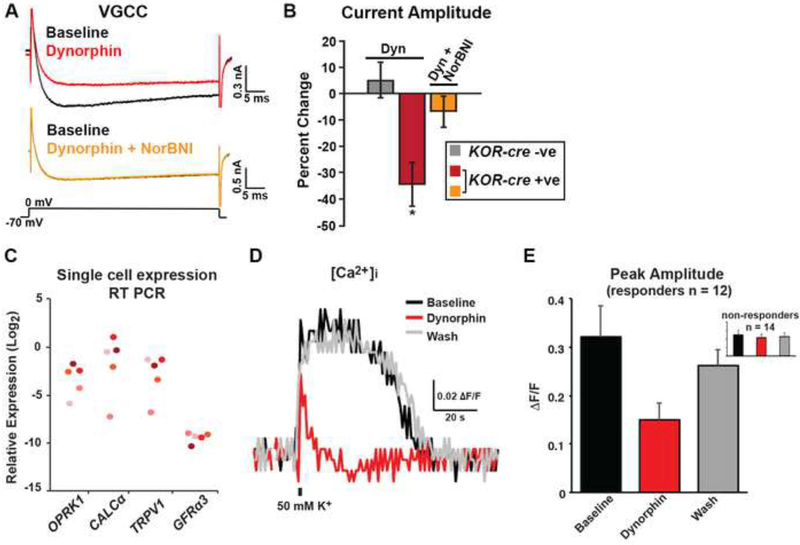
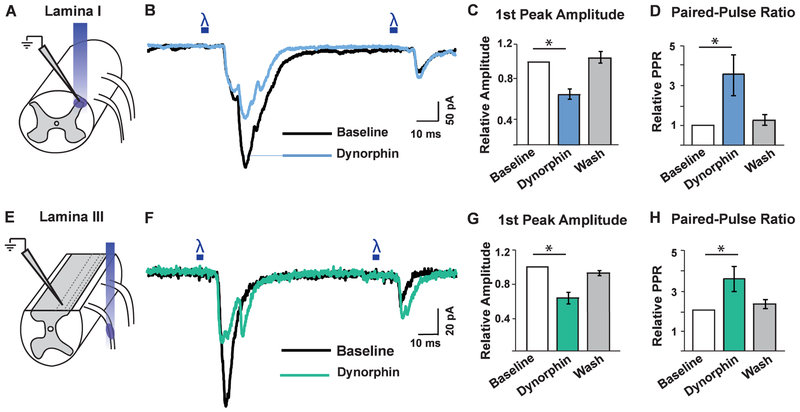
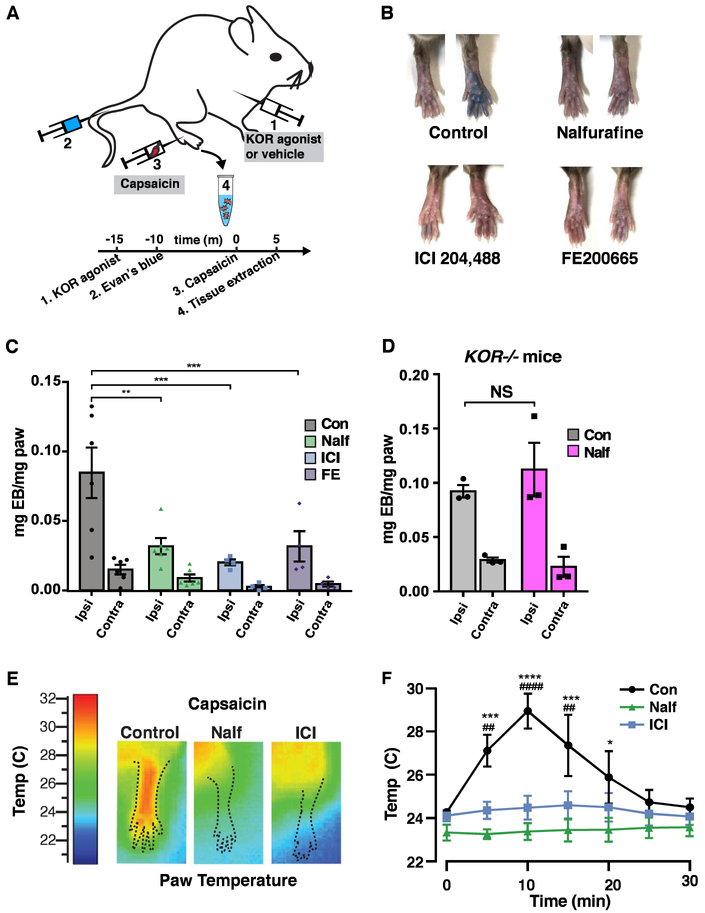
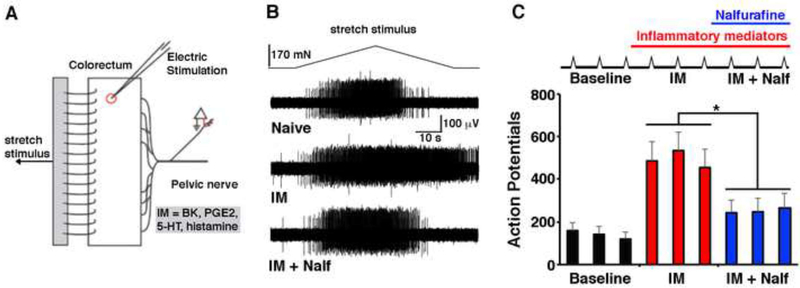
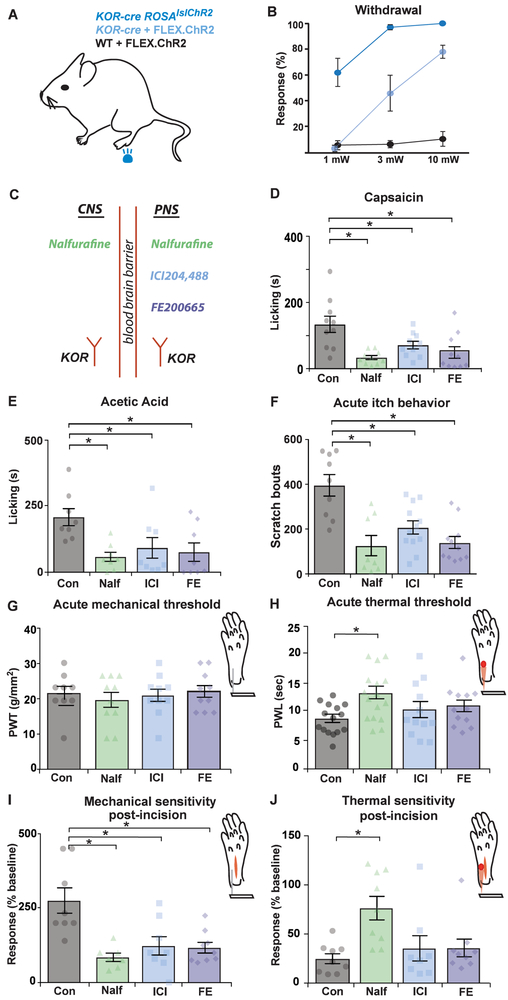
Primary afferents are known to be inhibited by kappa opioid receptor (KOR) signaling. However, the specific types of somatosensory neurons that express KOR remain unclear. Here, using a newly developed KOR-cre knockin allele, viral tracing, single-cell RT-PCR, and ex vivo recordings, we show that KOR is expressed in several populations of primary afferents: a subset of peptidergic sensory neurons, as well as low-threshold mechanoreceptors that form lanceolate or circumferential endings around hair follicles. We find that KOR acts centrally to inhibit excitatory neurotransmission from KOR-cre afferents in laminae I and III, and this effect is likely due to KOR-mediated inhibition of Ca2+ influx, which we observed in sensory neurons from both mouse and human. In the periphery, KOR signaling inhibits neurogenic inflammation and nociceptor sensitization by inflammatory mediators. Finally, peripherally restricted KOR agonists selectively reduce pain and itch behaviors, as well as mechanical hypersensitivity associated with a surgical incision. These experiments provide a rationale for the use of peripherally restricted KOR agonists for therapeutic treatment.
Keywords: DRG; Oprk1; dorsal root ganglia; dynorphin; human; itch; mouse; nalfurafine; pain; primary afferents.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
CONFLICTS OF INTEREST
None of the authors has a conflict of interest.
Figures
Comment in
-
Peripheral Kappa Opioid Receptor Signaling Takes on a Central Role.Neuron. 2018 Sep 19;99(6):1102-1104. doi: 10.1016/j.neuron.2018.09.006. Neuron. 2018. PMID: 30236278
References
-
- Bailey CP, and Connor M (2005). Opioids: cellular mechanisms of tolerance and physical dependence. Curr Opin Pharmacol 5, 60–68. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 NS096705/NS/NINDS NIH HHS/United States
- R01 DK107966/DK/NIDDK NIH HHS/United States
- R01 AR069951/AR/NIAMS NIH HHS/United States
- T32 NS073548/NS/NINDS NIH HHS/United States
- T32 NS007433/NS/NINDS NIH HHS/United States
- R01 DA044481/DA/NIDA NIH HHS/United States
- K99 DA031777/DA/NIDA NIH HHS/United States
- R01 DA034975/DA/NIDA NIH HHS/United States
- R01 NS023725/NS/NINDS NIH HHS/United States
- R00 DA031777/DA/NIDA NIH HHS/United States
- F31 NS092146/NS/NINDS NIH HHS/United States
- R01 NS083347/NS/NINDS NIH HHS/United States
- R01 DA041809/DA/NIDA NIH HHS/United States
- R01 AR063772/AR/NIAMS NIH HHS/United States
- R01 AR071720/AR/NIAMS NIH HHS/United States
- T32 MH020016/MH/NIMH NIH HHS/United States
- R01 AR060744/AR/NIAMS NIH HHS/United States
- R56 NS023725/NS/NINDS NIH HHS/United States
- F30 NS096860/NS/NINDS NIH HHS/United States
- R21 AR064445/AR/NIAMS NIH HHS/United States
- OT2 OD023859/OD/NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
