

The properties of Msh2-Msh6 ATP binding mutants suggest a signal amplification mechanism in DNA mismatch repair
- PMID: 30237169
- PMCID: PMC6254361
- DOI: 10.1074/jbc.RA118.005439
The properties of Msh2-Msh6 ATP binding mutants suggest a signal amplification mechanism in DNA mismatch repair
Abstract
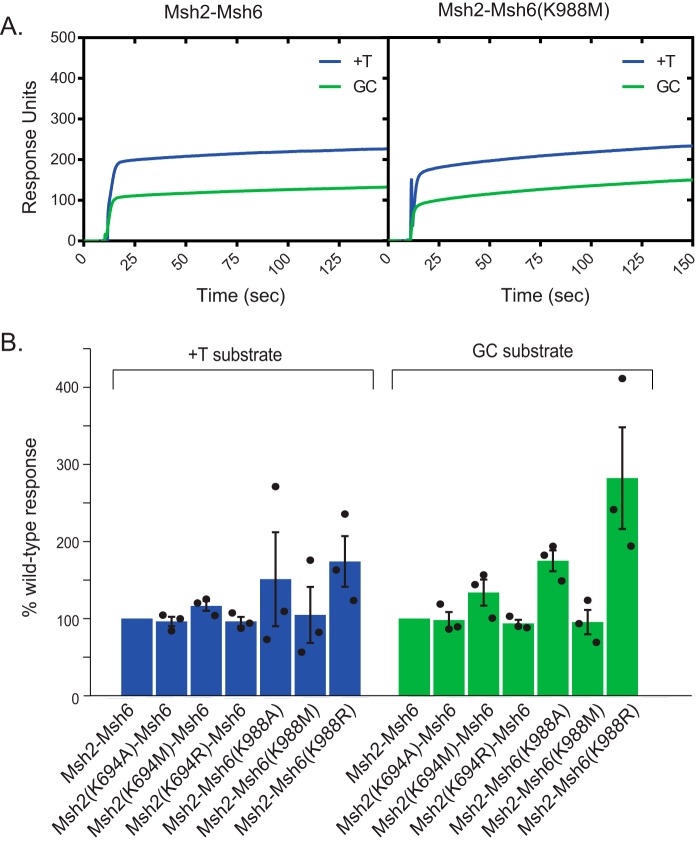
DNA mismatch repair (MMR) corrects mispaired DNA bases and small insertion/deletion loops generated by DNA replication errors. After binding a mispair, the eukaryotic mispair recognition complex Msh2-Msh6 binds ATP in both of its nucleotide-binding sites, which induces a conformational change resulting in the formation of an Msh2-Msh6 sliding clamp that releases from the mispair and slides freely along the DNA. However, the roles that Msh2-Msh6 sliding clamps play in MMR remain poorly understood. Here, using Saccharomyces cerevisiae, we created Msh2 and Msh6 Walker A nucleotide-binding site mutants that have defects in ATP binding in one or both nucleotide-binding sites of the Msh2-Msh6 heterodimer. We found that these mutations cause a complete MMR defect in vivo The mutant Msh2-Msh6 complexes exhibited normal mispair recognition and were proficient at recruiting the MMR endonuclease Mlh1-Pms1 to mispaired DNA. At physiological (2.5 mm) ATP concentration, the mutant complexes displayed modest partial defects in supporting MMR in reconstituted Mlh1-Pms1-independent and Mlh1-Pms1-dependent MMR reactions in vitro and in activation of the Mlh1-Pms1 endonuclease and showed a more severe defect at low (0.1 mm) ATP concentration. In contrast, five of the mutants were completely defective and one was mostly defective for sliding clamp formation at high and low ATP concentrations. These findings suggest that mispair-dependent sliding clamp formation triggers binding of additional Msh2-Msh6 complexes and that further recruitment of additional downstream MMR proteins is required for signal amplification of mispair binding during MMR.
Keywords: DNA binding protein; DNA endonuclease; DNA mismatch repair; DNA repair; DNA replication; Mlh1-Pms1; Msh2-Msh6; MutS; S. cerevisiae; exonuclease 1; mutagenesis.
© 2018 Graham et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures






Similar articles
-
Mispair-specific recruitment of the Mlh1-Pms1 complex identifies repair substrates of the Saccharomyces cerevisiae Msh2-Msh3 complex.J Biol Chem. 2014 Mar 28;289(13):9352-64. doi: 10.1074/jbc.M114.552190. Epub 2014 Feb 18. J Biol Chem. 2014. PMID: 24550389 Free PMC article.
-
Mlh1 interacts with both Msh2 and Msh6 for recruitment during mismatch repair.DNA Repair (Amst). 2022 Nov;119:103405. doi: 10.1016/j.dnarep.2022.103405. Epub 2022 Sep 14. DNA Repair (Amst). 2022. PMID: 36122480 Free PMC article.
-
Activation of Saccharomyces cerevisiae Mlh1-Pms1 Endonuclease in a Reconstituted Mismatch Repair System.J Biol Chem. 2015 Aug 28;290(35):21580-90. doi: 10.1074/jbc.M115.662189. Epub 2015 Jul 13. J Biol Chem. 2015. PMID: 26170454 Free PMC article.
-
DNA binding properties of the yeast Msh2-Msh6 and Mlh1-Pms1 heterodimers.Biol Chem. 2002 Jun;383(6):969-75. doi: 10.1515/BC.2002.103. Biol Chem. 2002. PMID: 12222686 Review.
-
DNA mismatch repair and mutation avoidance pathways.J Cell Physiol. 2002 Apr;191(1):28-41. doi: 10.1002/jcp.10077. J Cell Physiol. 2002. PMID: 11920679 Review.
Cited by
-
An integrative pan-cancer analysis reveals the oncogenic role of mutS homolog 6 (MSH6) in human tumors.Aging (Albany NY). 2021 Dec 7;13(23):25271-25290. doi: 10.18632/aging.203745. Epub 2021 Dec 7. Aging (Albany NY). 2021. PMID: 34941572 Free PMC article.
-
DeMAG predicts the effects of variants in clinically actionable genes by integrating structural and evolutionary epistatic features.Nat Commun. 2023 Apr 19;14(1):2230. doi: 10.1038/s41467-023-37661-z. Nat Commun. 2023. PMID: 37076482 Free PMC article.
-
The unstructured linker of Mlh1 contains a motif required for endonuclease function which is mutated in cancers.Proc Natl Acad Sci U S A. 2022 Oct 18;119(42):e2212870119. doi: 10.1073/pnas.2212870119. Epub 2022 Oct 10. Proc Natl Acad Sci U S A. 2022. PMID: 36215471 Free PMC article.
-
An antisense Alu transposon insertion/deletion polymorphism of ALDH1A1 may functionally associate with Parkinson's disease.BMC Geriatr. 2022 May 16;22(1):427. doi: 10.1186/s12877-022-03132-1. BMC Geriatr. 2022. PMID: 35578164 Free PMC article.
-
Comprehensive classification of ABC ATPases and their functional radiation in nucleoprotein dynamics and biological conflict systems.Nucleic Acids Res. 2020 Oct 9;48(18):10045-10075. doi: 10.1093/nar/gkaa726. Nucleic Acids Res. 2020. PMID: 32894288 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
