

Evidence for the Regulation of Gynoecium Morphogenesis by ETTIN via Cell Wall Dynamics
- PMID: 30237208
- PMCID: PMC6236608
- DOI: 10.1104/pp.18.00745
Evidence for the Regulation of Gynoecium Morphogenesis by ETTIN via Cell Wall Dynamics
Abstract
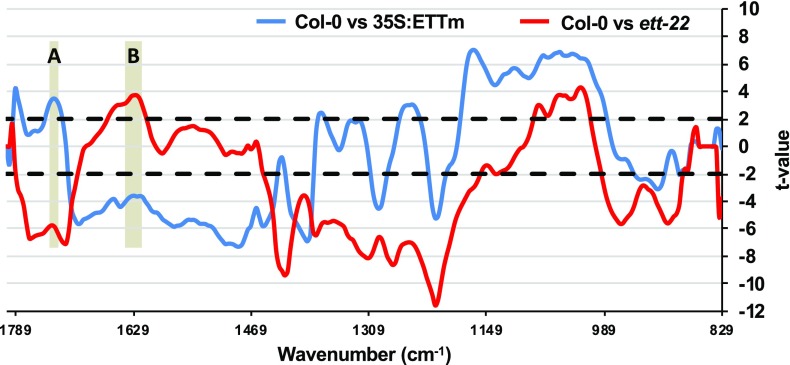
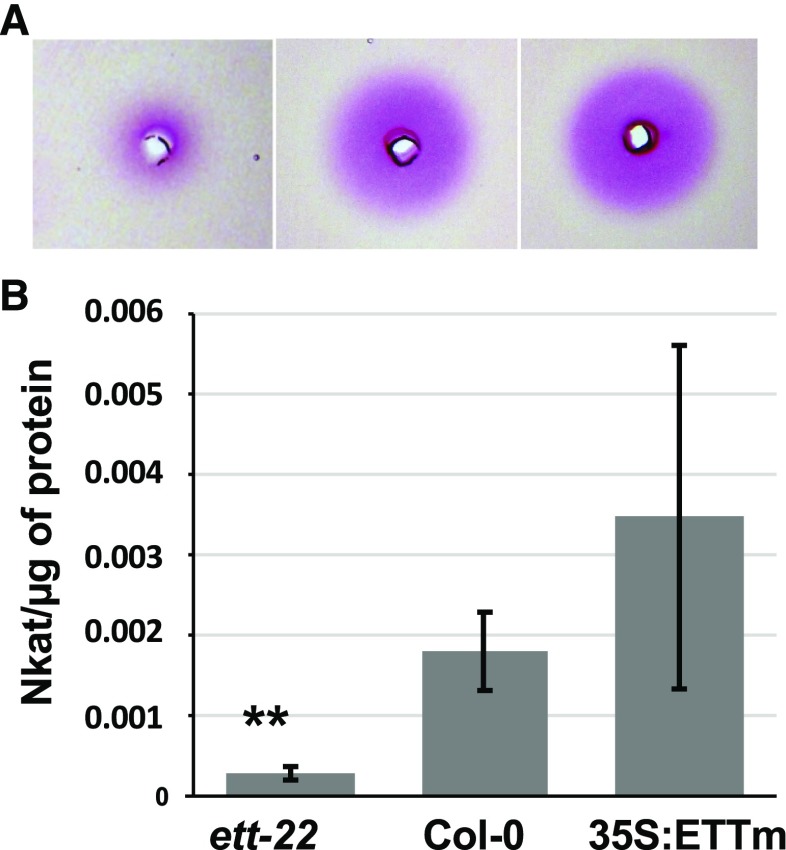
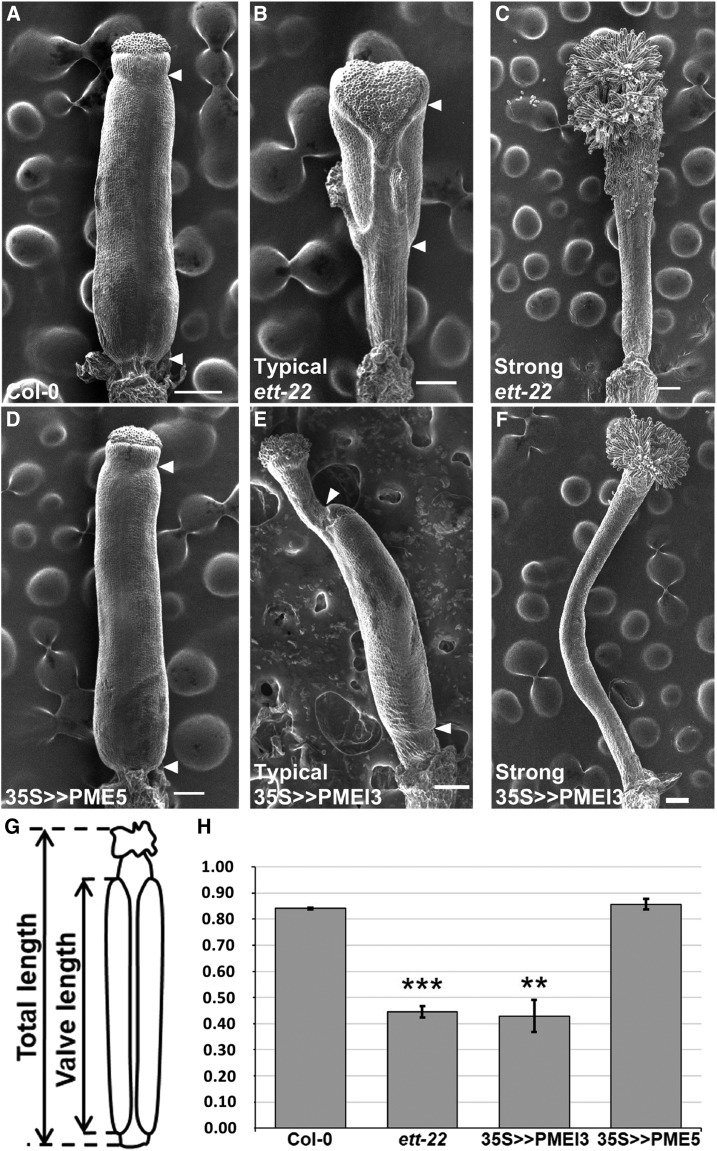
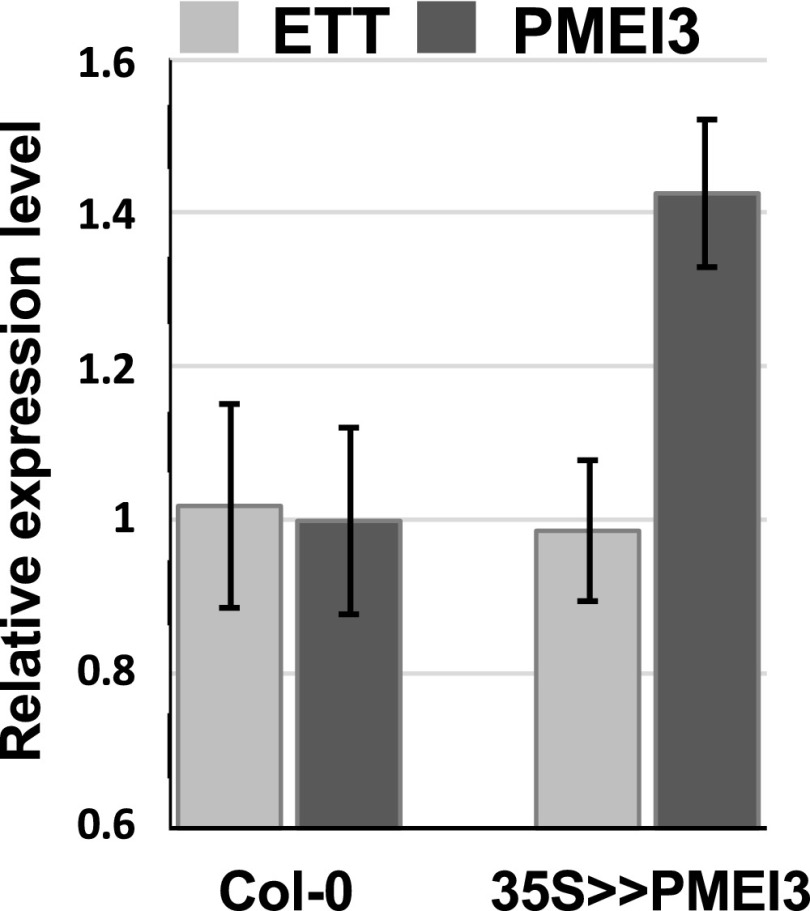
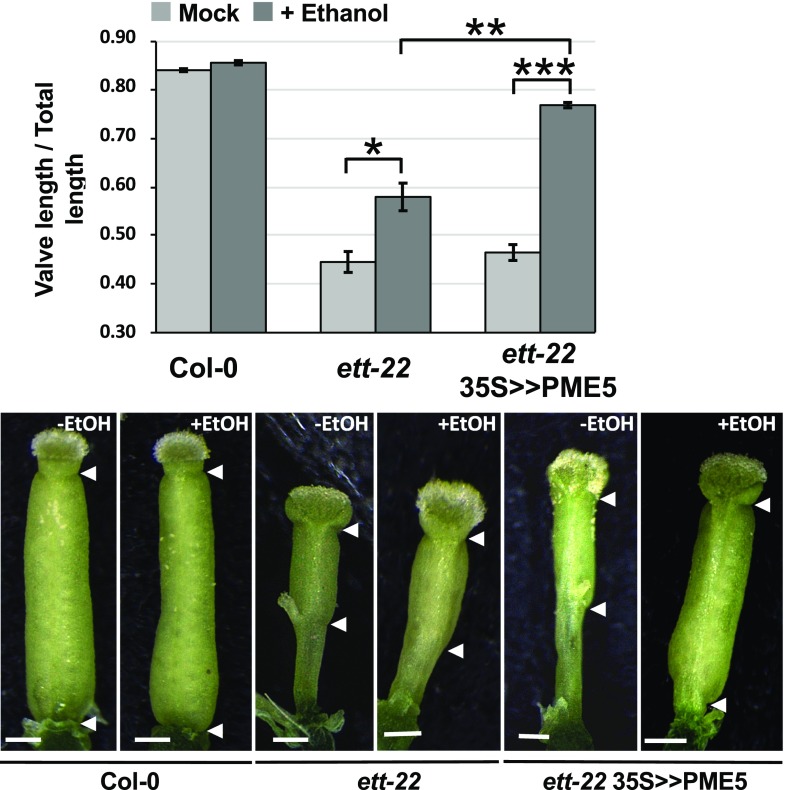
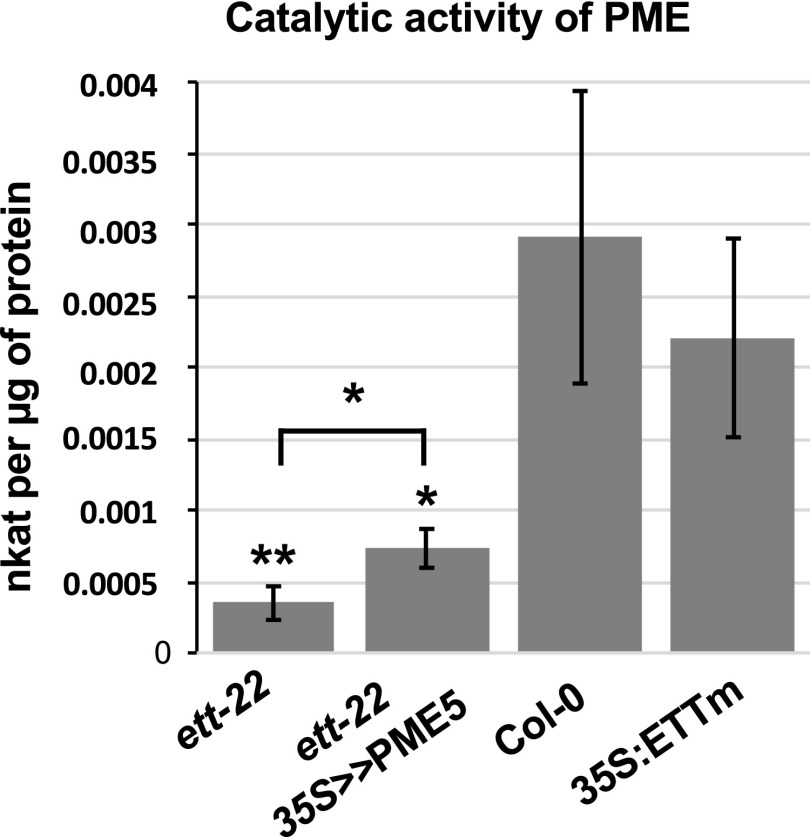
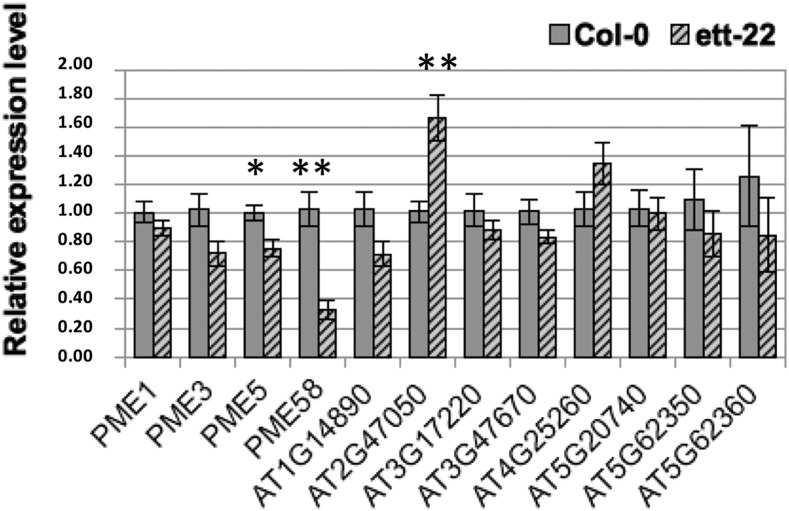
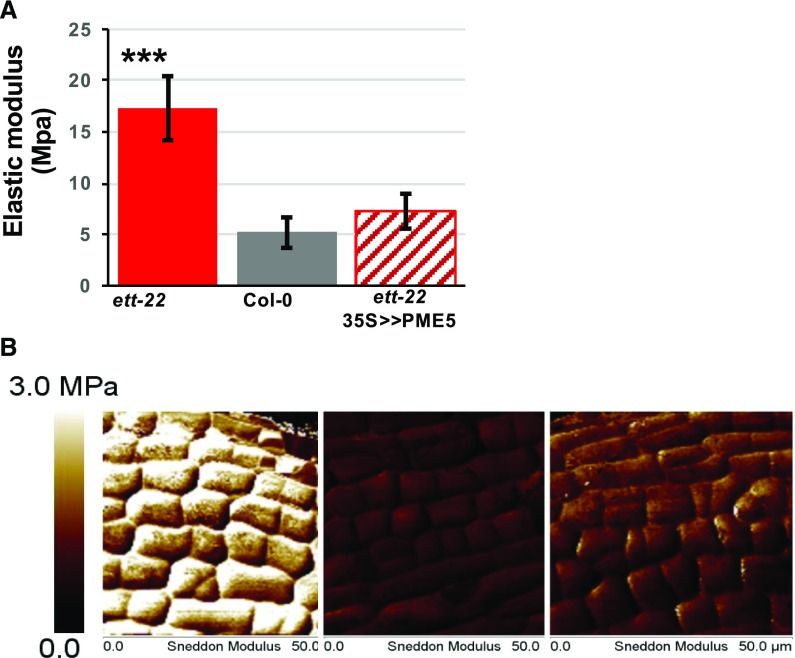
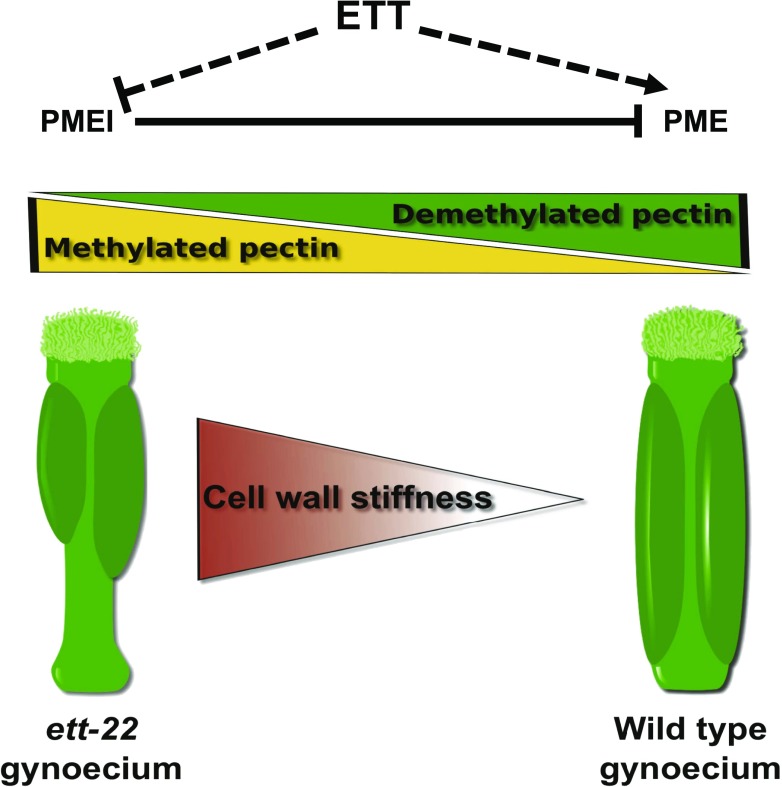
ETTIN (ETT) is an atypical member of the AUXIN RESPONSE FACTOR family of transcription factors that plays a crucial role in tissue patterning in the Arabidopsis (Arabidopsis thaliana) gynoecium. Though recent insights have provided valuable information on ETT's interactions with other components of auxin signaling, the biophysical mechanisms linking ETT to its ultimate effects on gynoecium morphology were until now unknown. Here, using techniques to assess cell-wall dynamics during gynoecium growth and development, we provide a coherent body of evidence to support a model in which ETT controls the elongation of the valve tissues of the gynoecium through the positive regulation of pectin methylesterase (PME) activity in the cell wall. This increase in PME activity results in an increase in the level of demethylesterified pectins and a consequent reduction in cell wall stiffness, leading to elongation of the valves. Though similar biophysical mechanisms have been shown to act in the stem apical meristem, leading to the expansion of organ primordia, our findings demonstrate that regulation of cell wall stiffness through the covalent modification of pectin also contributes to tissue patterning within a developing plant organ.
© 2018 American Society of Plant Biologists. All rights reserved.
Figures
References
-
- Asnacios A, Hamant O (2012) The mechanics behind cell polarity. Trends Cell Biol 22: 584–591 - PubMed
-
- Boutté Y, Crosnier M-T, Carraro N, Traas J, Satiat-Jeunemaitre B (2006) The plasma membrane recycling pathway and cell polarity in plants: studies on PIN proteins. J Cell Sci 119: 1255–1265 - PubMed
-
- Das P, Ito T, Wellmer F, Vernoux T, Dedieu A, Traas J, Meyerowitz EM (2009) Floral stem cell termination involves the direct regulation of AGAMOUS by PERIANTHIA. Development 136: 1605–1611 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
