

A gut-brain neural circuit for nutrient sensory transduction
- PMID: 30237325
- PMCID: PMC6417812
- DOI: 10.1126/science.aat5236
A gut-brain neural circuit for nutrient sensory transduction
Abstract
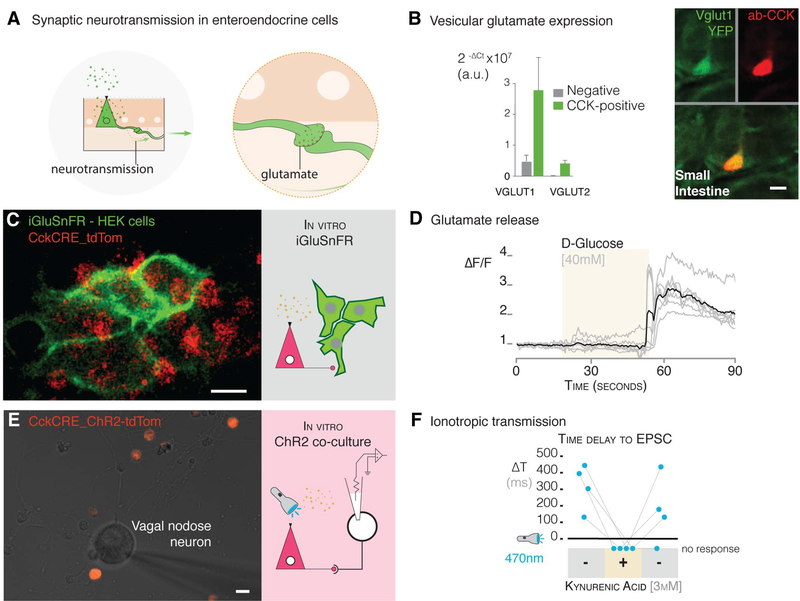
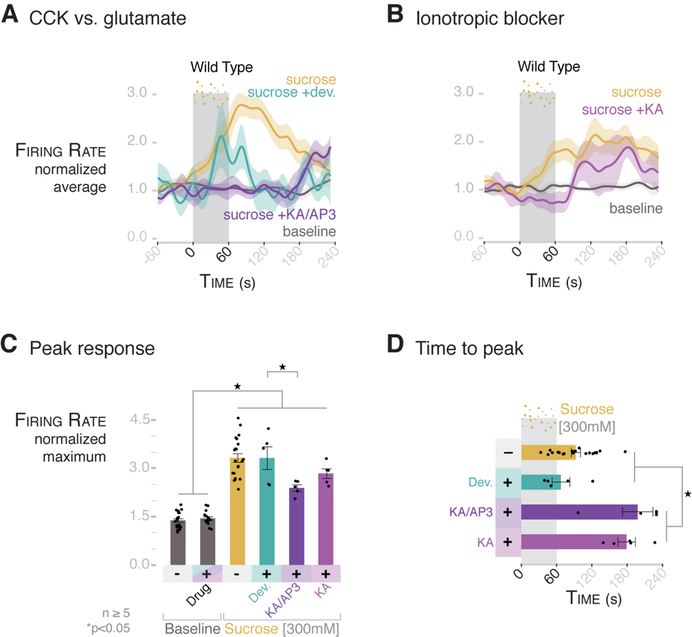
The brain is thought to sense gut stimuli only via the passive release of hormones. This is because no connection has been described between the vagus and the putative gut epithelial sensor cell-the enteroendocrine cell. However, these electrically excitable cells contain several features of epithelial transducers. Using a mouse model, we found that enteroendocrine cells synapse with vagal neurons to transduce gut luminal signals in milliseconds by using glutamate as a neurotransmitter. These synaptically connected enteroendocrine cells are referred to henceforth as neuropod cells. The neuroepithelial circuit they form connects the intestinal lumen to the brainstem in one synapse, opening a physical conduit for the brain to sense gut stimuli with the temporal precision and topographical resolution of a synapse.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
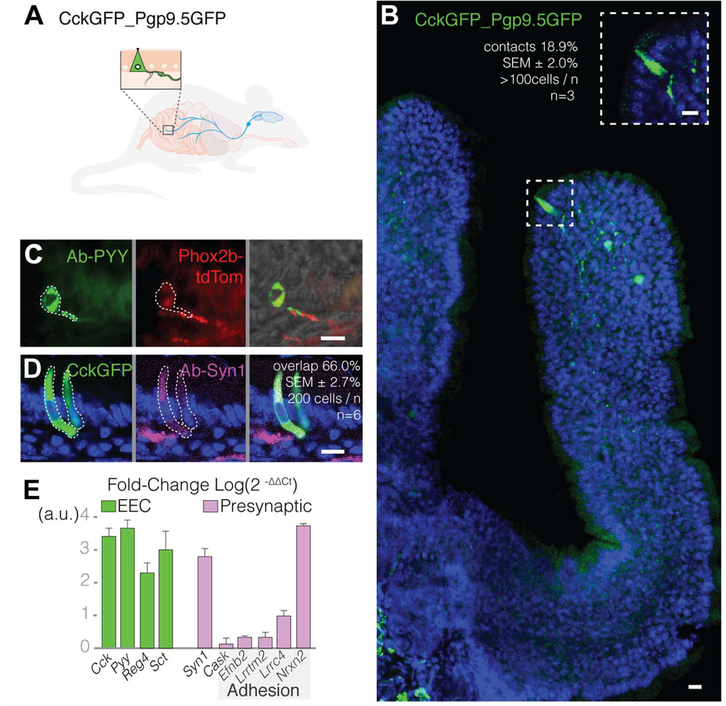
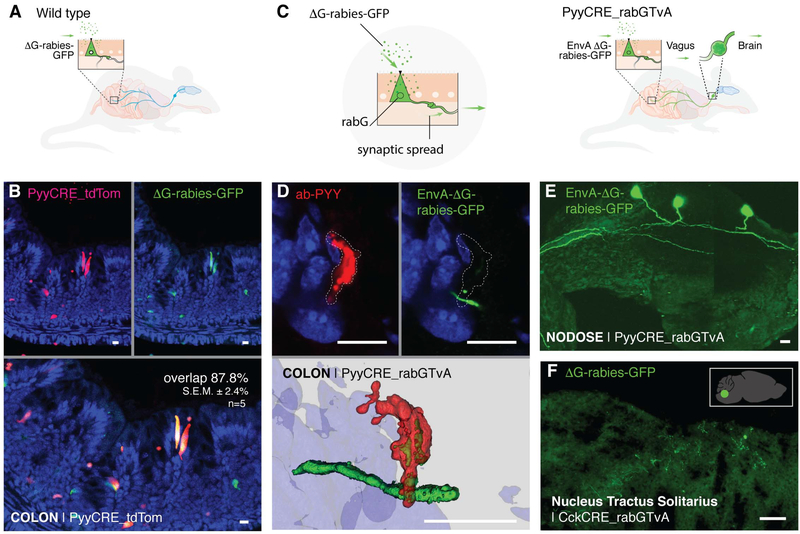
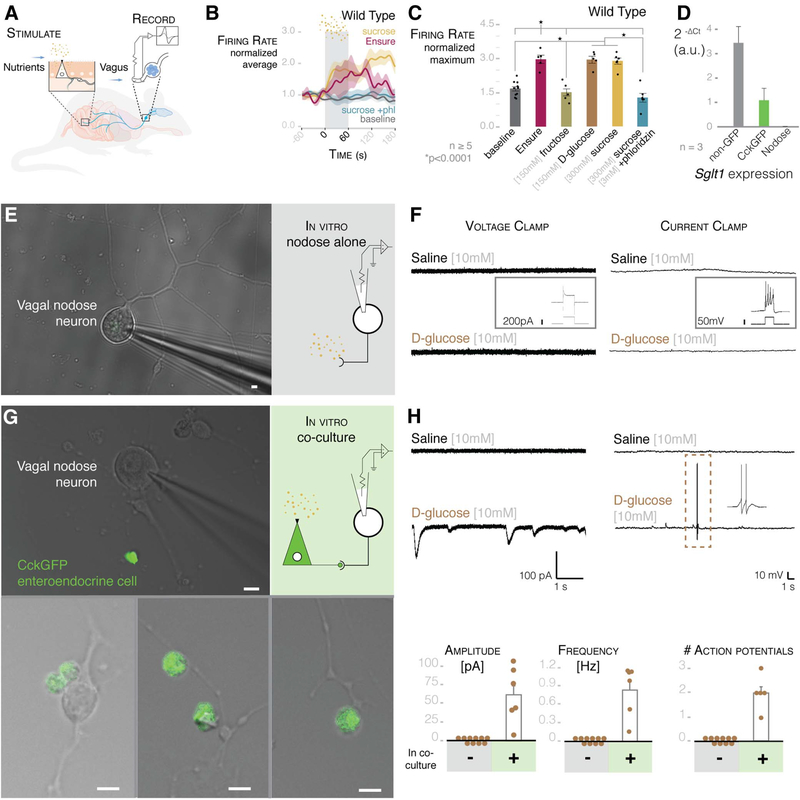
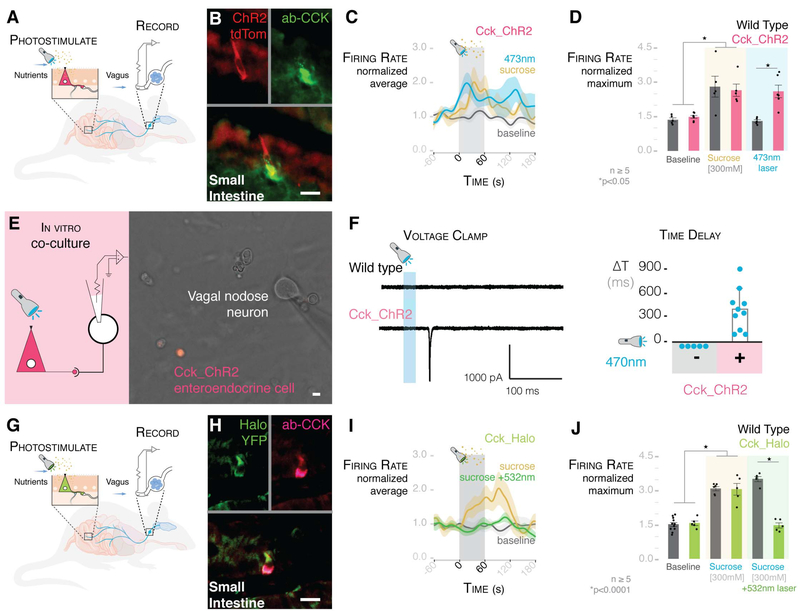
Comment in
-
A gut feeling.Science. 2018 Sep 21;361(6408):1203-1204. doi: 10.1126/science.aau9973. Science. 2018. PMID: 30237346 No abstract available.
-
A gut-brain neural connection for rapid nutrient sensing.Nat Rev Gastroenterol Hepatol. 2018 Nov;15(11):655. doi: 10.1038/s41575-018-0077-7. Nat Rev Gastroenterol Hepatol. 2018. PMID: 30323317 No abstract available.
-
[A gut-brain neural circuit for nutrient sensory transduction].Z Gastroenterol. 2019 Mar;57(3):335. doi: 10.1055/a-0821-1865. Epub 2019 Mar 12. Z Gastroenterol. 2019. PMID: 30861558 German. No abstract available.
References
-
- Alberts B, Bray D, Lewis J, Raff M, Roberts K, Watson JD, Molecular Biology of the Cell (Garland, ed. 3, 1994), pp. 907–982.
-
- Feyrter F, Über diffuse endokrine epitheliale Organe (J. A. Barth, Leipzig, Germany, 1938).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
