

Transcriptional profiling of Zygosaccharomyces bailii early response to acetic acid or copper stress mediated by ZbHaa1
- PMID: 30237501
- PMCID: PMC6147978
- DOI: 10.1038/s41598-018-32266-9
Transcriptional profiling of Zygosaccharomyces bailii early response to acetic acid or copper stress mediated by ZbHaa1
Abstract
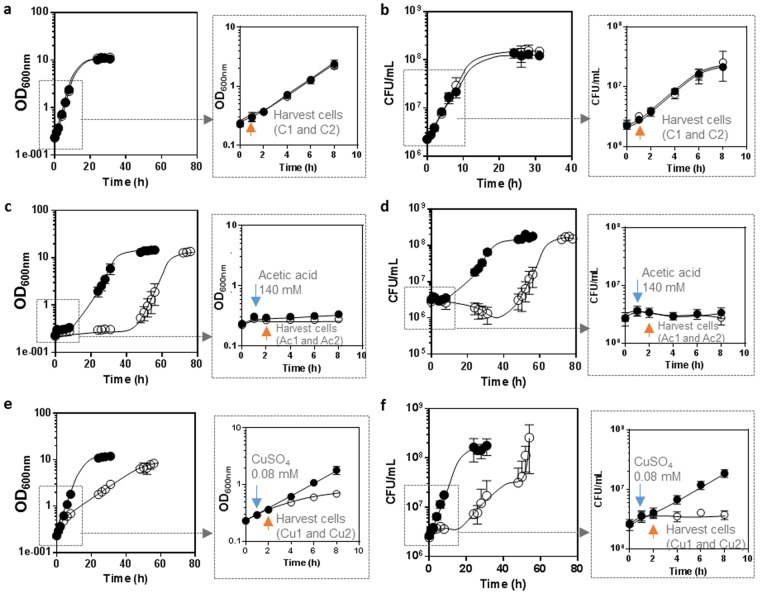
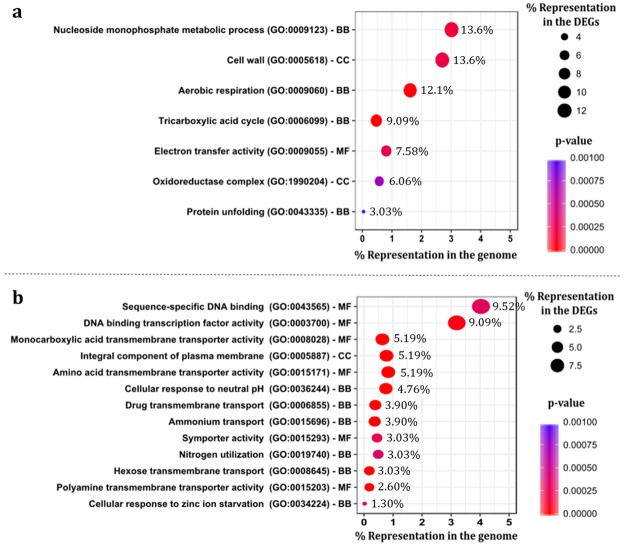
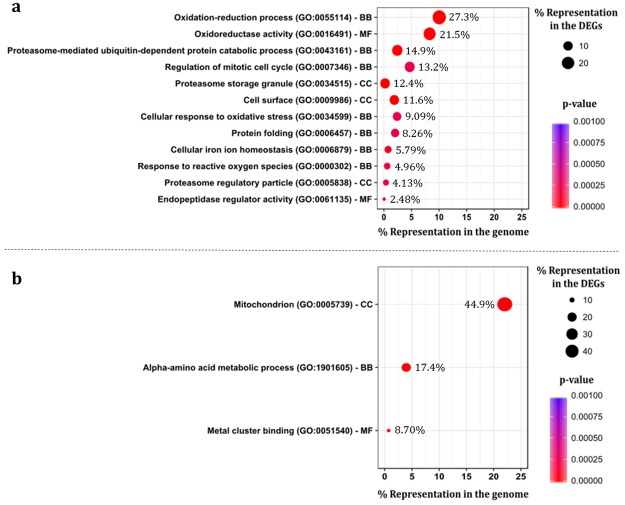
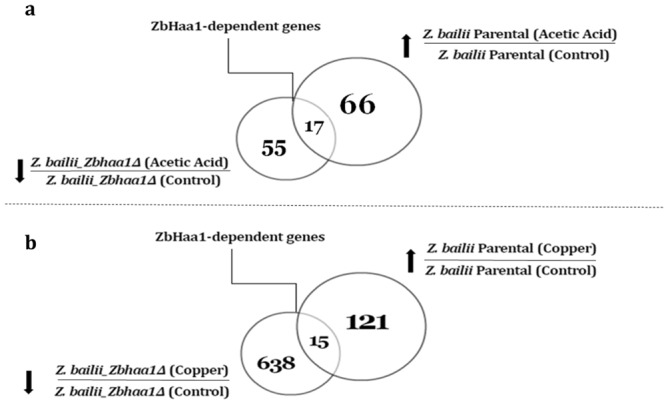
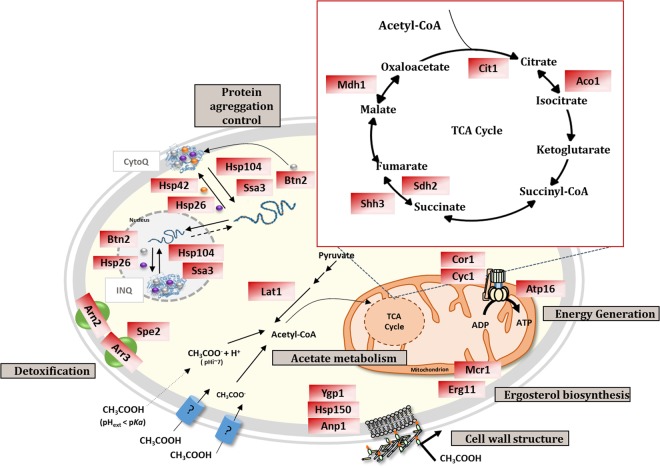
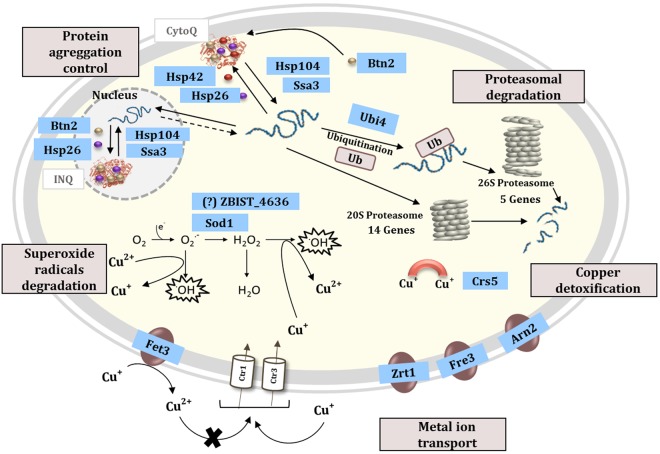
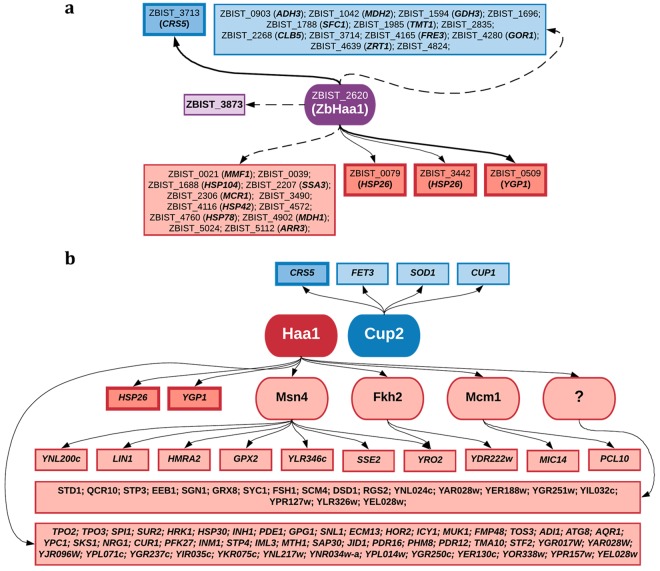
The non-conventional yeast species Zygosaccharomyces bailii is remarkably tolerant to acetic acid, a highly important microbial inhibitory compound in Food Industry and Biotechnology. ZbHaa1 is the functional homologue of S. cerevisiae Haa1 and a bifunctional transcription factor able to modulate Z. bailii adaptive response to acetic acid and copper stress. In this study, RNA-Seq was used to investigate genomic transcription changes in Z. bailii during early response to sublethal concentrations of acetic acid (140 mM, pH 4.0) or copper (0.08 mM) and uncover the regulatory network activated by these stresses under ZbHaa1 control. Differentially expressed genes in response to acetic acid exposure (297) are mainly related with the tricarboxylic acid cycle, protein folding and stabilization and modulation of plasma membrane composition and cell wall architecture, 17 of which, directly or indirectly, ZbHaa1-dependent. Copper stress induced the differential expression of 190 genes mainly involved in the response to oxidative stress, 15 ZbHaa1-dependent. This study provides valuable mechanistic insights regarding Z. bailii adaptation to acetic acid or copper stress, as well as useful information on transcription regulatory networks in pre-whole genome duplication (WGD) (Z. bailii) and post-WGD (S. cerevisiae) yeast species, contributing to the understanding of transcriptional networks' evolution in yeasts.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Sá-Correia, I., Guerreiro, J. F., Loureiro-Dias, M. C., Leão, C. & Côrte-Real, M. In Encyclopedia of Food Microbiology (eds Batt, C. A. & Tortorello, M. L.) 849–855 (Elsevier Ltd, Academic Press, 2014).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
