

COPII vesicles can affect the activity of antisense oligonucleotides by facilitating the release of oligonucleotides from endocytic pathways
- PMID: 30239896
- PMCID: PMC6212795
- DOI: 10.1093/nar/gky841
COPII vesicles can affect the activity of antisense oligonucleotides by facilitating the release of oligonucleotides from endocytic pathways
Abstract
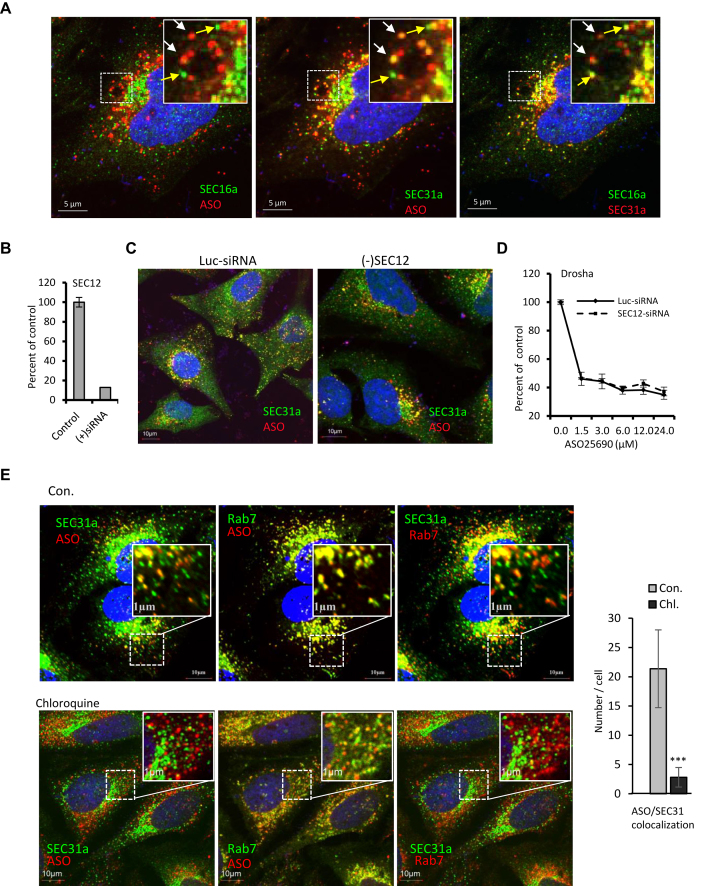
RNase H1-dependent, phosphorothioate-modified antisense oligonucleotides (PS-ASOs) can enter cells through endocytic pathways and need to be released from the membrane-enclosed organelles, a limiting step for antisense activity. Accumulating evidence has suggested that productive PS-ASO release mainly occurs from late endosomes (LEs). However, how PS-ASOs escape from LEs is not well understood. Here, we report that upon PS-ASO incubation, COPII vesicles, normally involved in ER-Golgi transport, can re-locate to PS-ASO-containing LEs. Reduction of COPII coat proteins significantly decreased PS-ASO activity, without affecting the levels of PS-ASO uptake and early-to-late endosome transport, but caused slower PS-ASO release from LEs. COPII co-localization with PS-ASOs at LEs does not require de novo assembly of COPII at ER. Interestingly, reduction of STX5 and P115, proteins involved in tethering and fusion of COPII vesicles with Golgi membranes, impaired COPII re-localization to LEs and decreased PS-ASO activity. STX5 can re-locate to LEs upon PS-ASO incubation, can bind PS-ASOs, and the binding appears to be required for this pathway. Our study reveals a novel release pathway in which PS-ASO incubation causes LE re-localization of STX5, which mediates the recruitment of COPII vesicles to LEs to facilitate endosomal PS-ASO release, and identifies another key PS-ASO binding protein.
Figures









References
-
- Dias N., Stein C.A.. Antisense oligonucleotides: basic concepts and mechanisms. Mol. Cancer Ther. 2002; 1:347–355. - PubMed
-
- Crooke S.T., Vickers T., Lima W.F., Wu H.-J.. Crooke ST. Mechanisms of Antisense Drug Action, an Introduction. Antisense Drug Technology: Principles, Strategies, and Applications. 2008; 2nd ednBoca Raton: CRC Press; 3–46.
-
- Crooke S.T., Witztum J.L., Bennett C.F., Baker B.F.. RNA-Targeted therapeutics. Cell metabolism. 2018; 27:714–739. - PubMed
-
- Bennett C.F., Baker B.F., Pham N., Swayze E., Geary R.S.. Pharmacology of antisense drugs. Annu. Rev. Pharmacol. Toxicol. 2017; 57:81–105. - PubMed
-
- Bennett C.F., Swayze E.E.. RNA targeting therapeutics: molecular mechanisms of antisense oligonucleotides as a therapeutic platform. Annu. Rev. Pharmacol. Toxicol. 2010; 50:259–293. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
