

Something special about CO-dependent CO2 fixation
- PMID: 30240136
- PMCID: PMC6282760
- DOI: 10.1111/febs.14664
Something special about CO-dependent CO2 fixation
Abstract
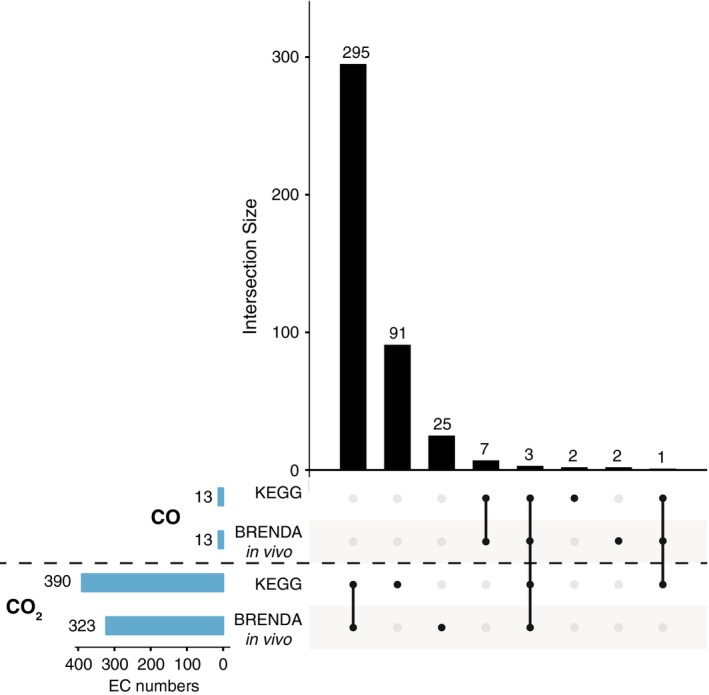
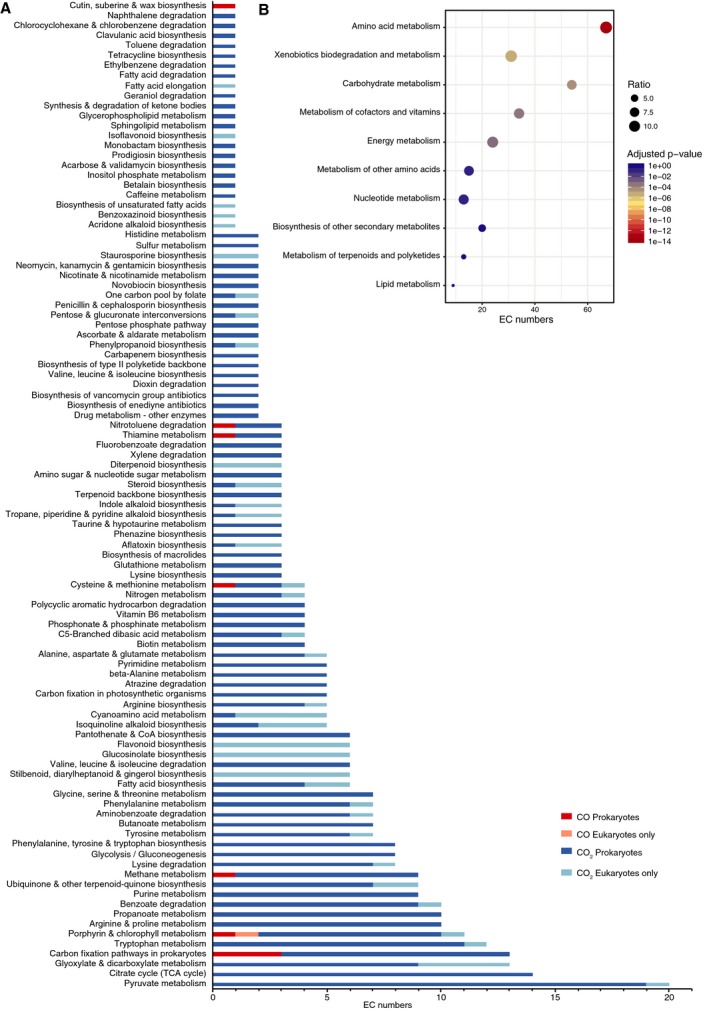
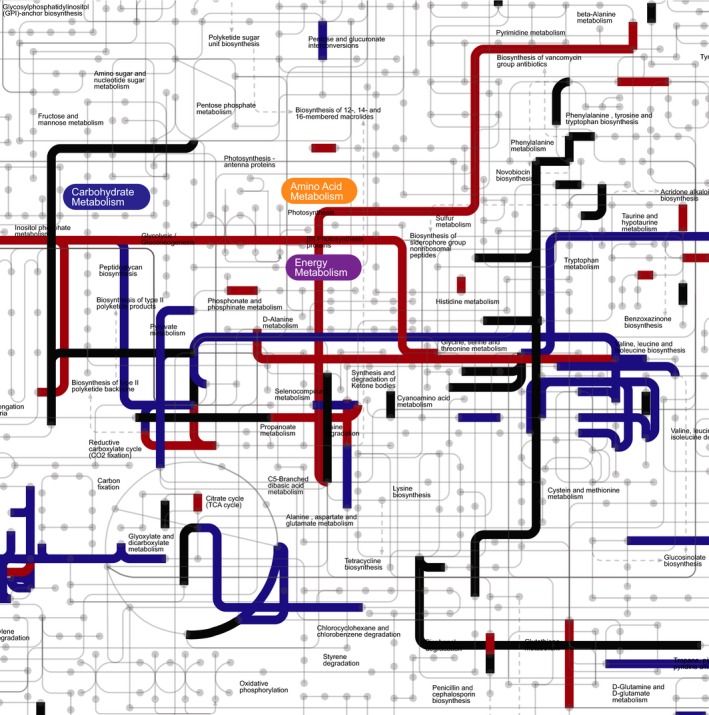
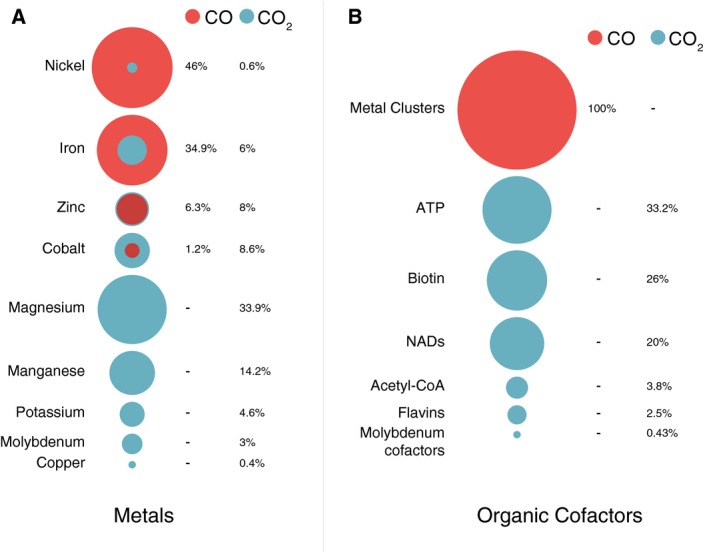
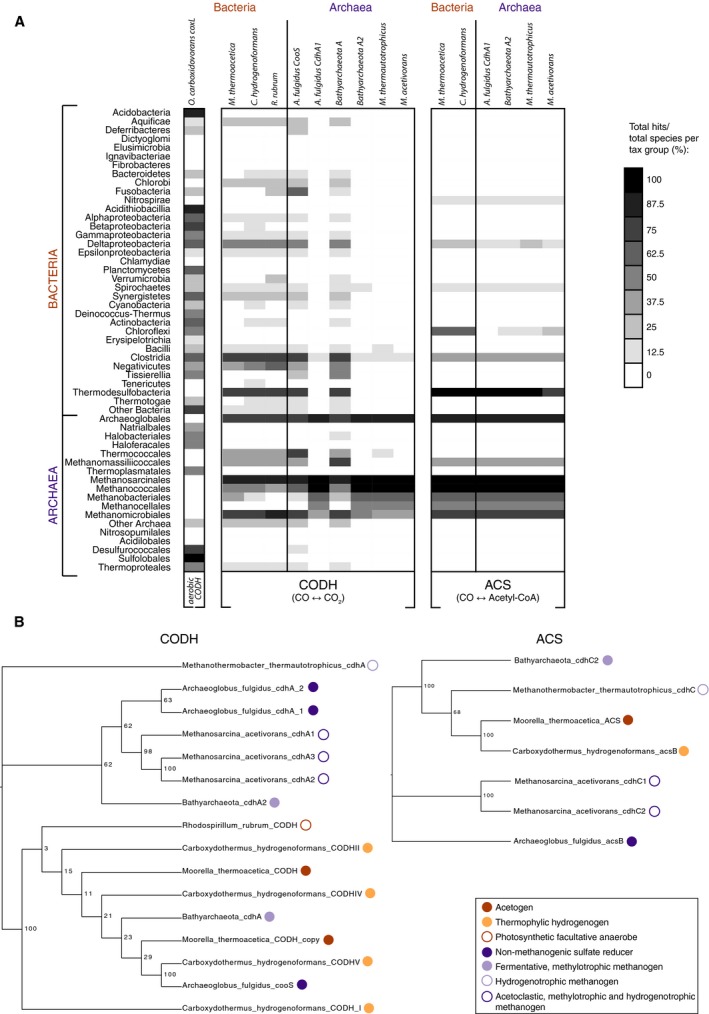
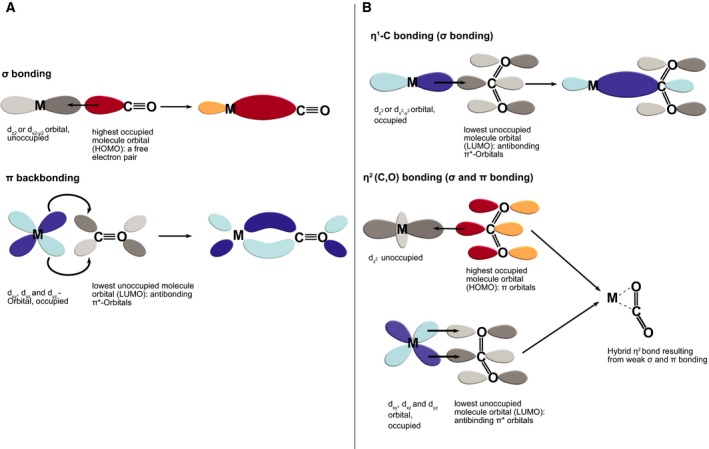
Carbon dioxide enters metabolism via six known CO2 fixation pathways, of which only one is linear, exergonic in the direction of CO2 -assimilation, and present in both bacterial and archaeal anaerobes - the Wood-Ljungdahl (WL) or reductive acetyl-CoA pathway. Carbon monoxide (CO) plays a central role in the WL pathway as an energy rich intermediate. Here, we scan the major biochemical reaction databases for reactions involving CO and CO2 . We identified 415 reactions corresponding to enzyme commission (EC) numbers involving CO2 , which are non-randomly distributed across different biochemical pathways. Their taxonomic distribution, reversibility under physiological conditions, cofactors and prosthetic groups are summarized. In contrast to CO2 , only 15 reaction classes involving CO were detected. Closer inspection reveals that CO interfaces with metabolism and the carbon cycle at only two enzymes: anaerobic carbon monoxide dehydrogenase (CODH), a Ni- and Fe-containing enzyme that generates CO for CO2 fixation in the WL pathway, and aerobic CODH, a Mo- and Cu-containing enzyme that oxidizes environmental CO as an electron source. The CO-dependent reaction of the WL pathway involves carbonyl insertion into a methyl carbon-nickel at the Ni-Fe-S A-cluster of acetyl-CoA synthase (ACS). It appears that no alternative mechanisms to the CO-dependent reaction of ACS have evolved in nearly 4 billion years, indicating an ancient and mechanistically essential role for CO at the onset of metabolism.
Keywords: CODH/ACS; carbon dioxide; carbon monoxide; enzymatic reactions; metabolic networks.
© 2018 The Authors. The FEBS Journal published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Figures
Similar articles
-
Heterologous Expression of the Clostridium carboxidivorans CO Dehydrogenase Alone or Together with the Acetyl Coenzyme A Synthase Enables both Reduction of CO2 and Oxidation of CO by Clostridium acetobutylicum.Appl Environ Microbiol. 2017 Aug 1;83(16):e00829-17. doi: 10.1128/AEM.00829-17. Print 2017 Aug 15. Appl Environ Microbiol. 2017. PMID: 28625981 Free PMC article.
-
Carbon monoxide. Toxic gas and fuel for anaerobes and aerobes: carbon monoxide dehydrogenases.Met Ions Life Sci. 2014;14:37-69. doi: 10.1007/978-94-017-9269-1_3. Met Ions Life Sci. 2014. PMID: 25416390 Review.
-
Insights into CO2 Fixation Pathway of Clostridium autoethanogenum by Targeted Mutagenesis.mBio. 2016 May 24;7(3):e00427-16. doi: 10.1128/mBio.00427-16. mBio. 2016. PMID: 27222467 Free PMC article.
-
Investigations of the efficient electrocatalytic interconversions of carbon dioxide and carbon monoxide by nickel-containing carbon monoxide dehydrogenases.Met Ions Life Sci. 2014;14:71-97. doi: 10.1007/978-94-017-9269-1_4. Met Ions Life Sci. 2014. PMID: 25416391 Free PMC article. Review.
-
Evolutionary history of carbon monoxide dehydrogenase/acetyl-CoA synthase, one of the oldest enzymatic complexes.Proc Natl Acad Sci U S A. 2018 Feb 6;115(6):E1166-E1173. doi: 10.1073/pnas.1716667115. Epub 2018 Jan 22. Proc Natl Acad Sci U S A. 2018. PMID: 29358391 Free PMC article.
Cited by
-
The Autotrophic Core: An Ancient Network of 404 Reactions Converts H2, CO2, and NH3 into Amino Acids, Bases, and Cofactors.Microorganisms. 2021 Feb 23;9(2):458. doi: 10.3390/microorganisms9020458. Microorganisms. 2021. PMID: 33672143 Free PMC article.
-
Catalysts, autocatalysis and the origin of metabolism.Interface Focus. 2019 Dec 6;9(6):20190072. doi: 10.1098/rsfs.2019.0072. Epub 2019 Oct 18. Interface Focus. 2019. PMID: 31641438 Free PMC article. Review.
-
The Biotic and Abiotic Carbon Monoxide Formation During Aerobic Co-digestion of Dairy Cattle Manure With Green Waste and Sawdust.Front Bioeng Biotechnol. 2019 Oct 29;7:283. doi: 10.3389/fbioe.2019.00283. eCollection 2019. Front Bioeng Biotechnol. 2019. PMID: 31737615 Free PMC article.
-
Time-Course Transcriptome of Parageobacillus thermoglucosidasius DSM 6285 Grown in the Presence of Carbon Monoxide and Air.Int J Mol Sci. 2020 May 29;21(11):3870. doi: 10.3390/ijms21113870. Int J Mol Sci. 2020. PMID: 32485888 Free PMC article.
-
Magnetic interactions between metal sites in complex enzymes.J Biol Inorg Chem. 2025 Aug;30(4-5):329-344. doi: 10.1007/s00775-025-02120-1. Epub 2025 Jul 24. J Biol Inorg Chem. 2025. PMID: 40705057 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
