

Identification of the Human Skeletal Stem Cell
- PMID: 30241615
- PMCID: PMC6400492
- DOI: 10.1016/j.cell.2018.07.029
Identification of the Human Skeletal Stem Cell
Abstract
Stem cell regulation and hierarchical organization of human skeletal progenitors remain largely unexplored. Here, we report the isolation of a self-renewing and multipotent human skeletal stem cell (hSSC) that generates progenitors of bone, cartilage, and stroma, but not fat. Self-renewing and multipotent hSSCs are present in fetal and adult bones and can also be derived from BMP2-treated human adipose stroma (B-HAS) and induced pluripotent stem cells (iPSCs). Gene expression analysis of individual hSSCs reveals overall similarity between hSSCs obtained from different sources and partially explains skewed differentiation toward cartilage in fetal and iPSC-derived hSSCs. hSSCs undergo local expansion in response to acute skeletal injury. In addition, hSSC-derived stroma can maintain human hematopoietic stem cells (hHSCs) in serum-free culture conditions. Finally, we combine gene expression and epigenetic data of mouse skeletal stem cells (mSSCs) and hSSCs to identify evolutionarily conserved and divergent pathways driving SSC-mediated skeletogenesis. VIDEO ABSTRACT.
Keywords: ATAC-sequencing; HSC; and stromal progenitor; bone; bone fracture repair; bone marrow niche; cartilage; human skeletal stem cell; single cell RNA-sequencing.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interests
Patent is pending for the isolation, derivation, and use of human skeletal stem cells and downstream skeletal progenitors.
Figures
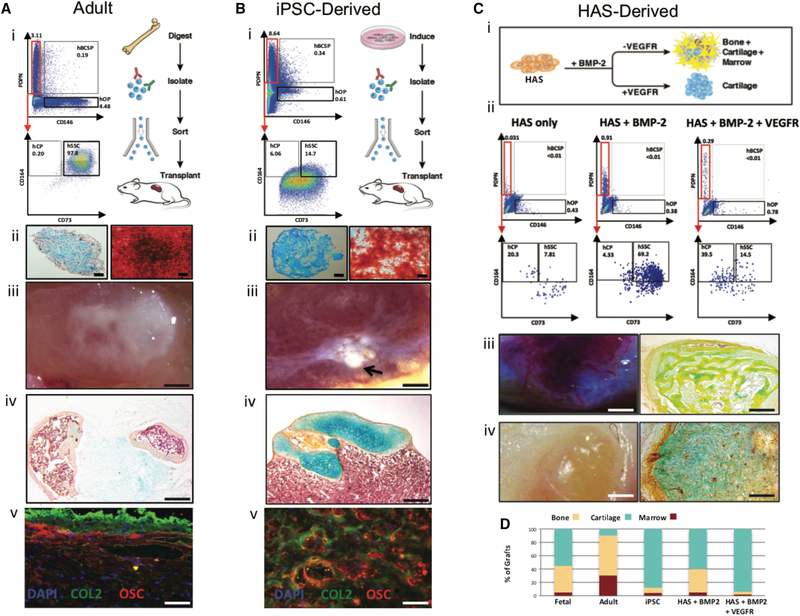
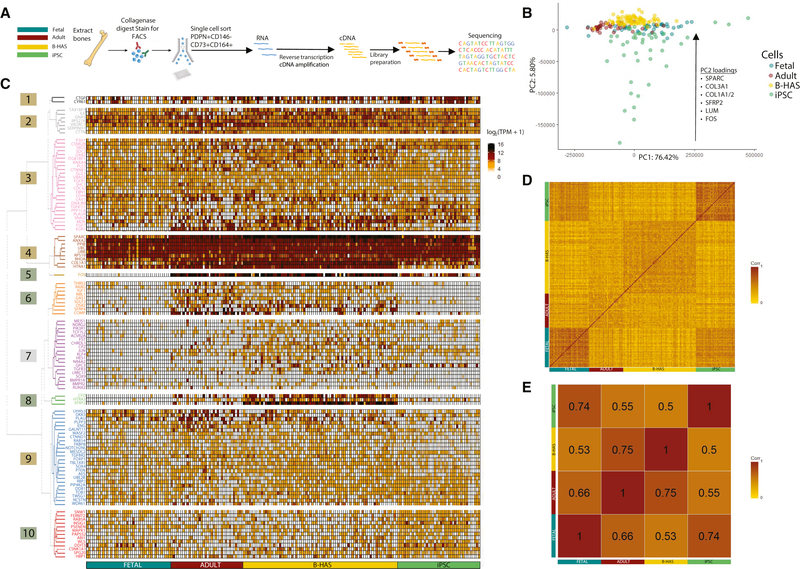
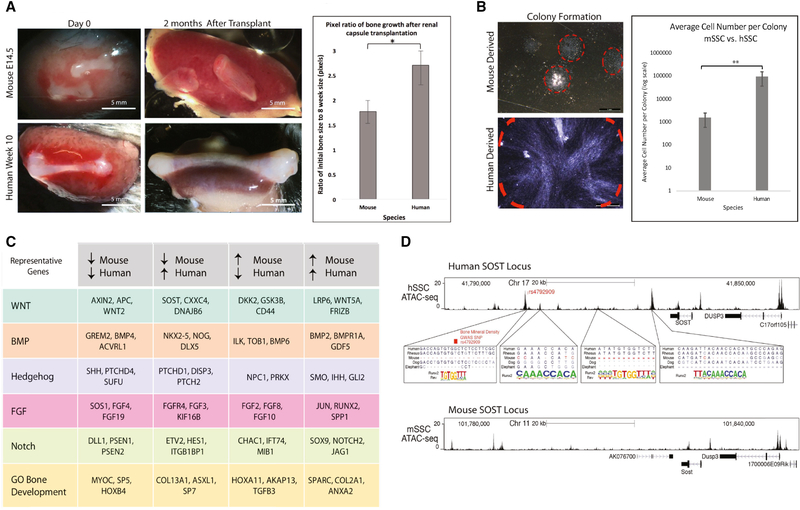
Comment in
-
Human Skeletal Stem Cells: The Markers Provide Some Clues in the Hunt for Hidden Treasure.Cell Stem Cell. 2018 Oct 4;23(4):462-463. doi: 10.1016/j.stem.2018.09.006. Cell Stem Cell. 2018. PMID: 30290174
-
Close to the bone - in search of the skeletal stem cell.Nat Rev Rheumatol. 2018 Dec;14(12):687-688. doi: 10.1038/s41584-018-0121-6. Nat Rev Rheumatol. 2018. PMID: 30425330 No abstract available.
References
-
- Bensidhoum M, Chapel A, Francois S, Demarquay C, Mazurier C, Fouillard L, Bouchet S, Bertho JM, Gourmelon P, Aigueperse J, et al. (2004). Homing of in vitro expanded Stro-1- or Stro-1+human mesenchymal stem cells into the NOD/SCID mouse and their role in supporting human CD34 cell engraftment. Blood 103, 3313–3319. - PubMed
-
- Beyth S, Schroeder J, and Liebergall M (2011). Stem cells in bone diseases: current clinical practice. Br Med Bull 99, 199–210. - PubMed
Publication types
MeSH terms
Grants and funding
- R21 DE019274/DE/NIDCR NIH HHS/United States
- R01 AR063717/AR/NIAMS NIH HHS/United States
- K99 AG049958/AG/NIA NIH HHS/United States
- R01 DE026730/DE/NIDCR NIH HHS/United States
- R56 DE025597/DE/NIDCR NIH HHS/United States
- R01 DE021683/DE/NIDCR NIH HHS/United States
- U01 HL099999/HL/NHLBI NIH HHS/United States
- U01 HL099776/HL/NHLBI NIH HHS/United States
- R21 DE024230/DE/NIDCR NIH HHS/United States
- R00 AG049958/AG/NIA NIH HHS/United States
- R01 DE027323/DE/NIDCR NIH HHS/United States
- P50 HG007735/HG/NHGRI NIH HHS/United States
- R01 AR055650/AR/NIAMS NIH HHS/United States
- U24 DE026914/DE/NIDCR NIH HHS/United States
- R01 DK115600/DK/NIDDK NIH HHS/United States
- R01 CA086065/CA/NCI NIH HHS/United States
- R01 HL058770/HL/NHLBI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
