

Dietary Fatty Acids Sustain the Growth of the Human Gut Microbiota
- PMID: 30242004
- PMCID: PMC6193386
- DOI: 10.1128/AEM.01525-18
Dietary Fatty Acids Sustain the Growth of the Human Gut Microbiota
Abstract
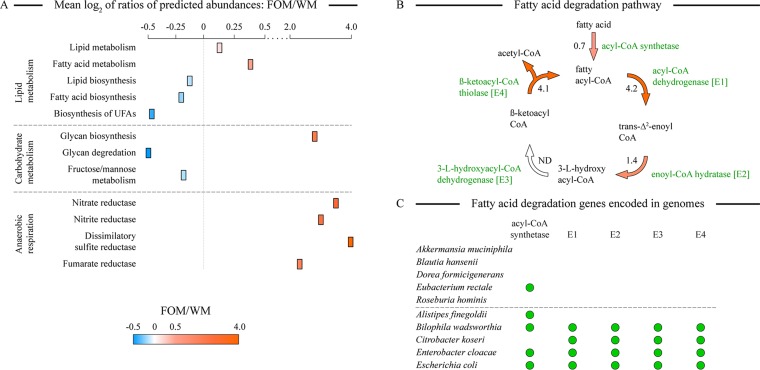
While a substantial amount of dietary fats escape absorption in the human small intestine and reach the colon, the ability of resident microbiota to utilize these dietary fats for growth has not been investigated in detail. In this study, we used an in vitro multivessel simulator system of the human colon to reveal that the human gut microbiota is able to utilize typically consumed dietary fatty acids to sustain growth. Gut microbiota adapted quickly to a macronutrient switch from a balanced Western diet-type medium to its variant lacking carbohydrates and proteins. We defined specific genera that increased in their abundances on the fats-only medium, including Alistipes, Bilophila, and several genera of the class Gammaproteobacteria In contrast, the abundances of well-known glycan and protein degraders, including Bacteroides, Clostridium, and Roseburia spp., were reduced under such conditions. The predicted prevalences of microbial genes coding for fatty acid degradation enzymes and anaerobic respiratory reductases were significantly increased in the fats-only environment, whereas the abundance of glycan degradation genes was diminished. These changes also resulted in lower microbial production of short-chain fatty acids and antioxidants. Our findings provide justification for the previously observed alterations in gut microbiota observed in human and animal studies of high-fat diets.IMPORTANCE Increased intake of fats in many developed countries has raised awareness of potentially harmful and beneficial effects of high fat consumption on human health. Some dietary fats escape digestion in the small intestine and reach the colon where they can be metabolized by gut microbiota. We show that human gut microbes are able to maintain a complex community when supplied with dietary fatty acids as the only nutrient and carbon sources. Such fatty acid-based growth leads to lower production of short-chain fatty acids and antioxidants by community members, which potentially have negative health consequences on the host.
Keywords: Western diet; dietary fats; microbial digestion; microbiota; nutrition.
Copyright © 2018 American Society for Microbiology.
Figures
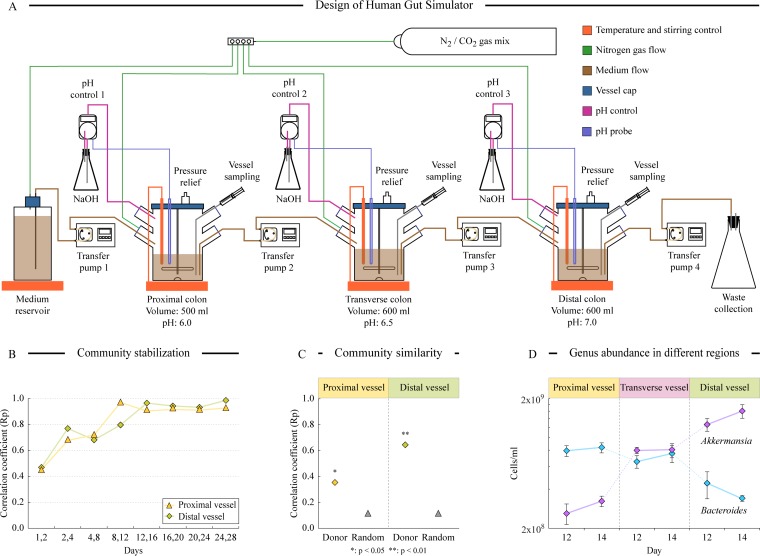
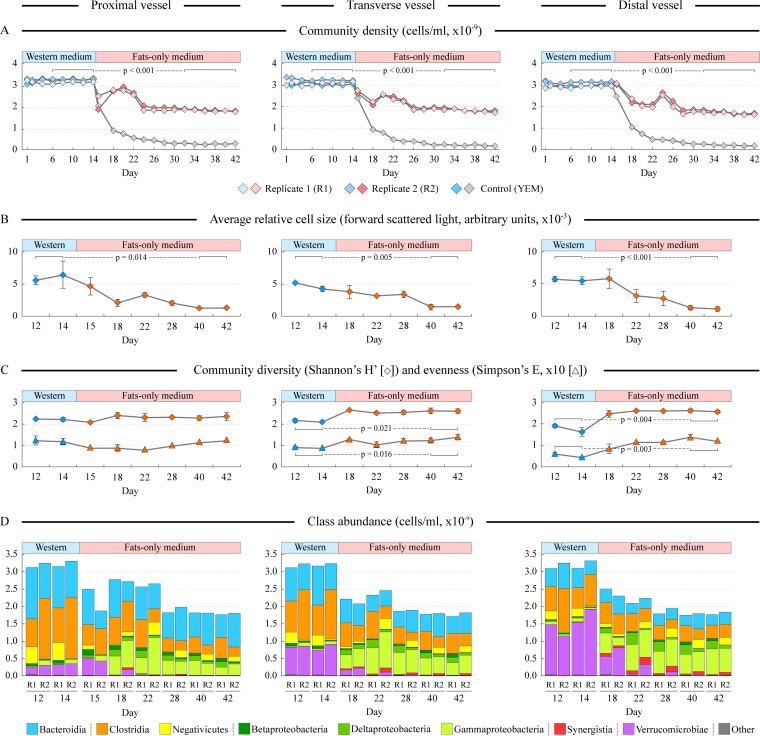
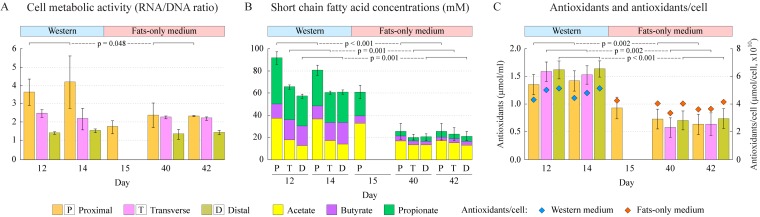
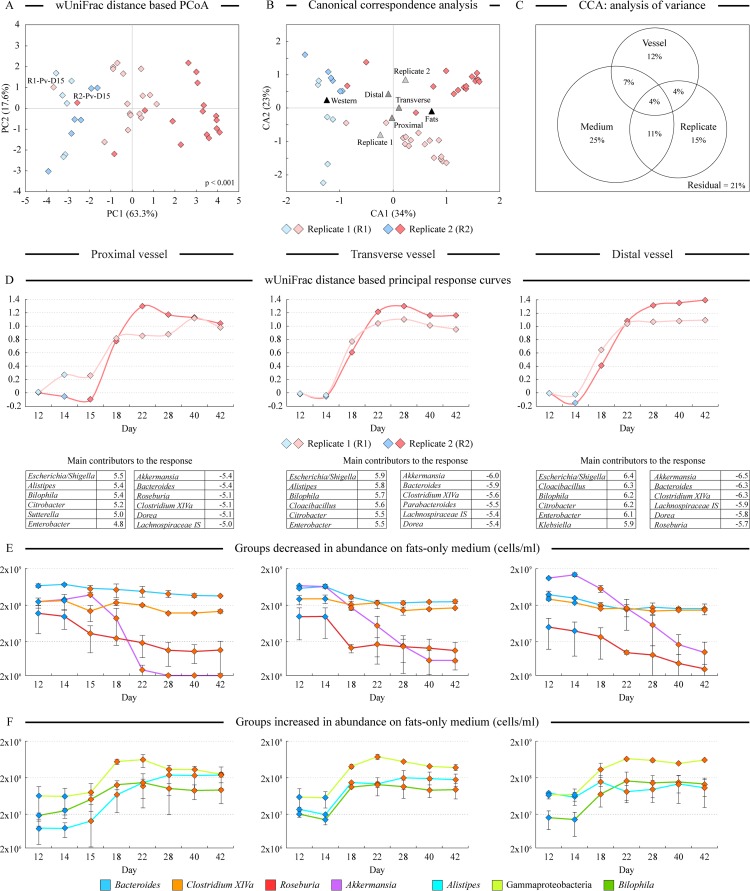
References
-
- Van Horn LV. 2010. Report of the dietary guidelines advisory committee on the dietary guidelines for Americans. U.S. Department of Agriculture, Washington, DC.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
