

Impaired Notch Signaling Leads to a Decrease in p53 Activity and Mitotic Catastrophe in Aged Muscle Stem Cells
- PMID: 30244867
- PMCID: PMC6173623
- DOI: 10.1016/j.stem.2018.08.019
Impaired Notch Signaling Leads to a Decrease in p53 Activity and Mitotic Catastrophe in Aged Muscle Stem Cells
Abstract
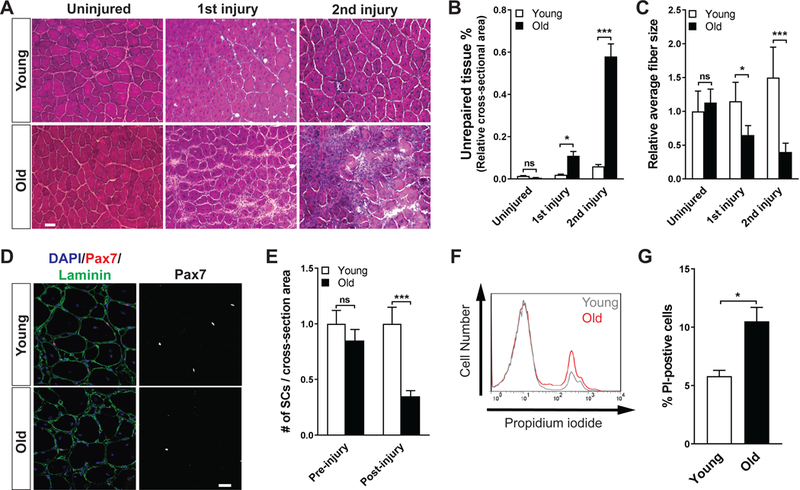
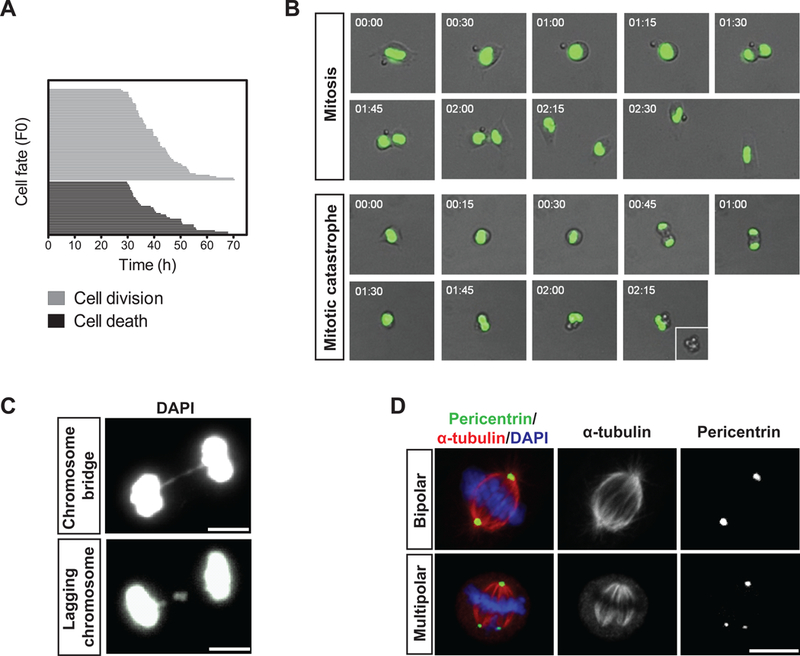
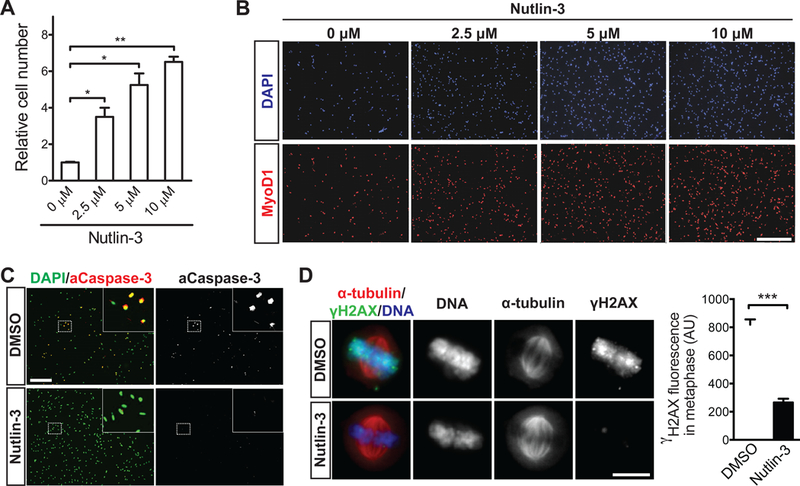
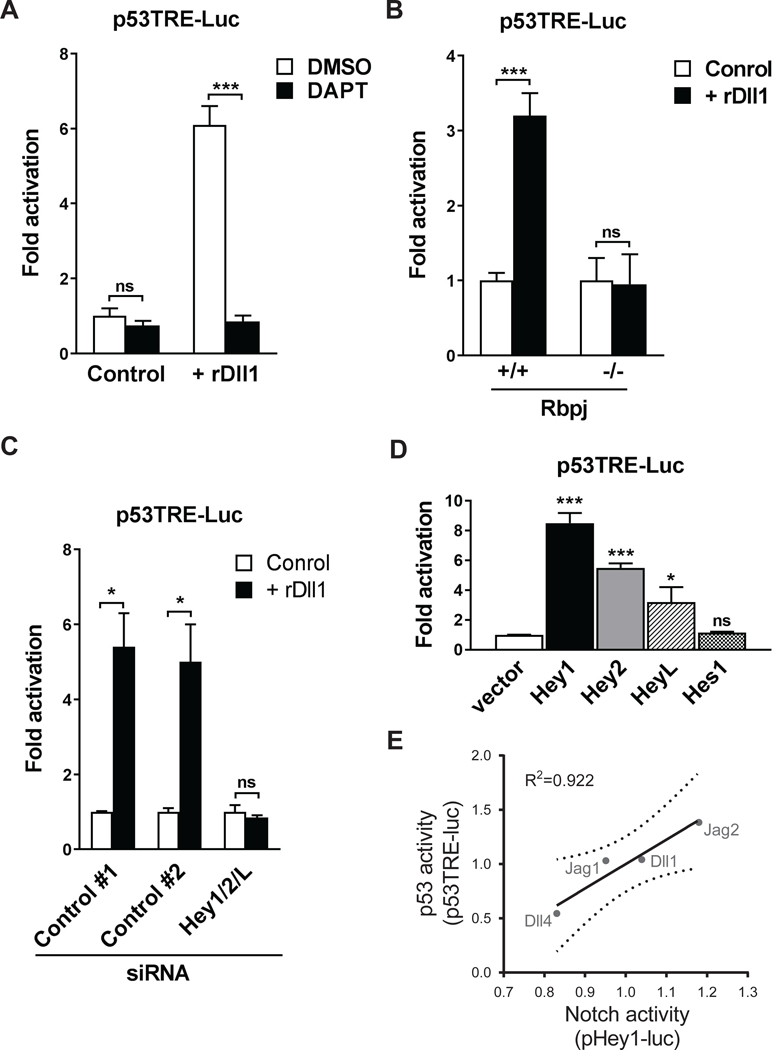
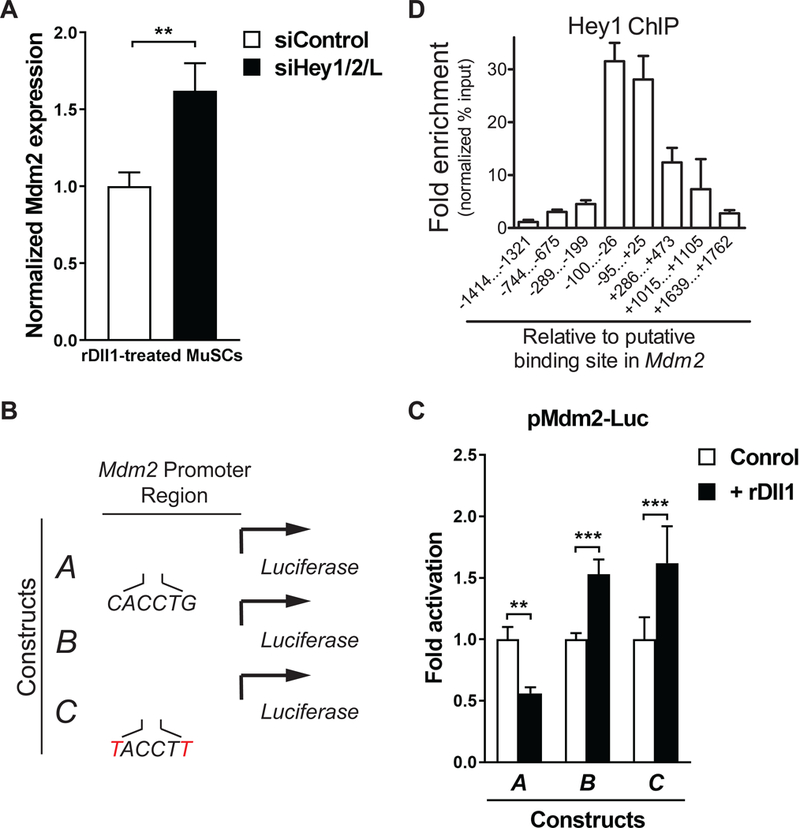
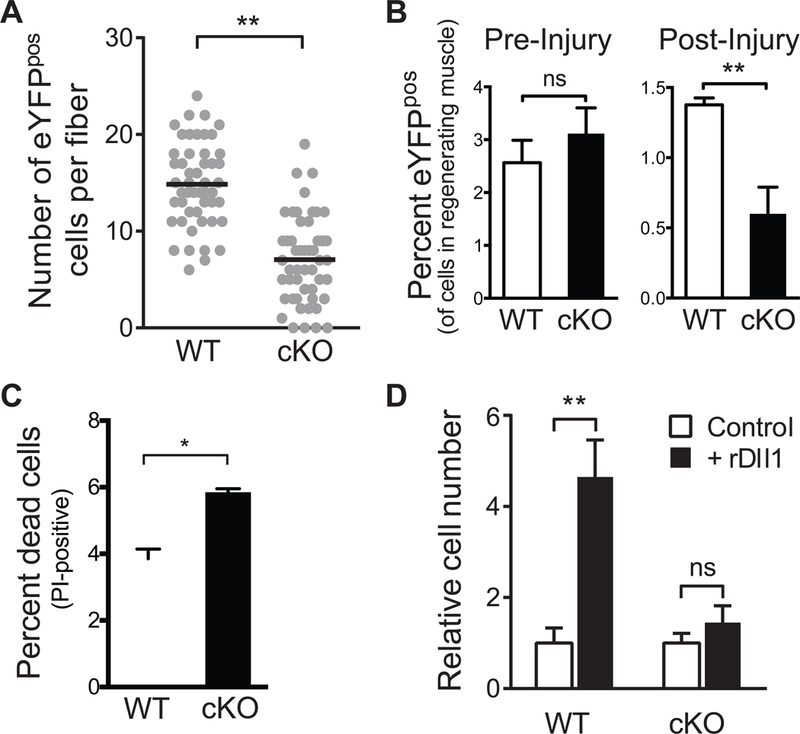
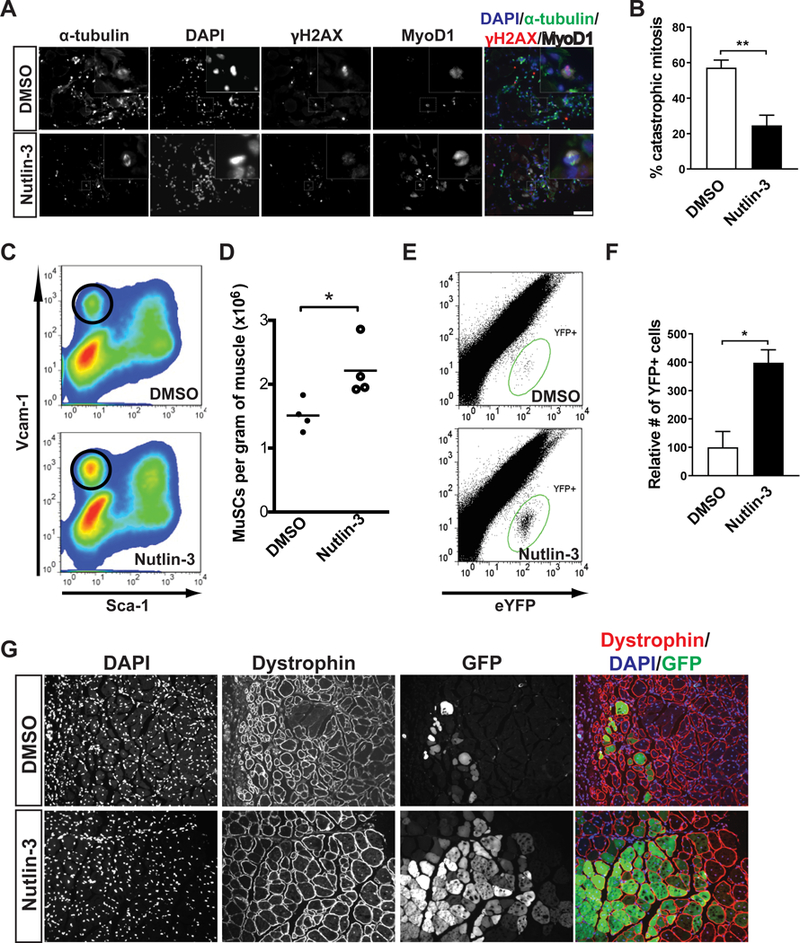
The decline of tissue regenerative potential with age correlates with impaired stem cell function. However, limited strategies are available for therapeutic modulation of stem cell function during aging. Using skeletal muscle stem cells (MuSCs) as a model system, we identify cell death by mitotic catastrophe as a cause of impaired stem cell proliferative expansion in aged animals. The mitotic cell death is caused by a deficiency in Notch activators in the microenvironment. We discover that ligand-dependent stimulation of Notch activates p53 in MuSCs via inhibition of Mdm2 expression through Hey transcription factors during normal muscle regeneration and that this pathway is impaired in aged animals. Pharmacologic activation of p53 promotes the expansion of aged MuSCs in vivo. Altogether, these findings illuminate a Notch-p53 signaling axis that plays an important role in MuSC survival during activation and is dysregulated during aging, contributing to the age-related decline in muscle regenerative potential.
Keywords: Notch; aging; mitotic catastrophe; muscle regeneration; muscle stem cells; p53; satellite cells; stem cells.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures
References
-
- Brack AS, Conboy MJ, Roy S, Lee M, Kuo CJ, Keller C, and Rando TA (2007). Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science 317, 807–810. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
