

Structural Basis of TRPV4 N Terminus Interaction with Syndapin/PACSIN1-3 and PIP2
- PMID: 30244966
- PMCID: PMC6281781
- DOI: 10.1016/j.str.2018.08.002
Structural Basis of TRPV4 N Terminus Interaction with Syndapin/PACSIN1-3 and PIP2
Abstract
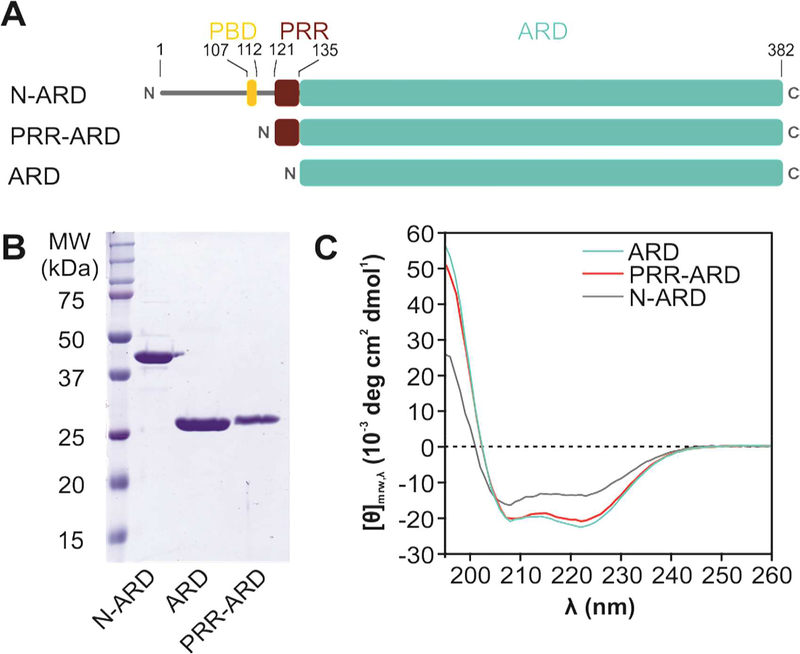
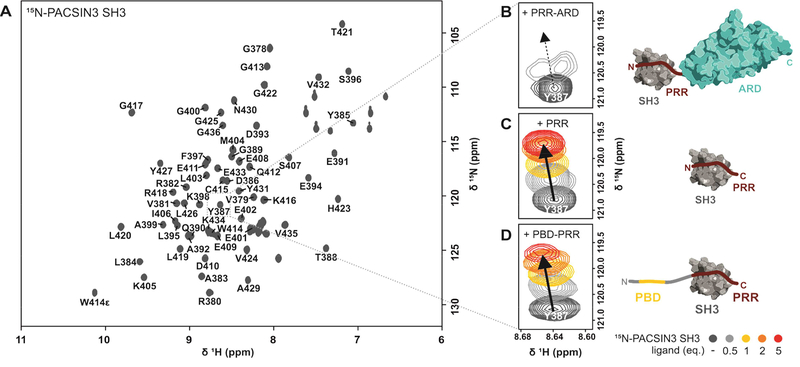
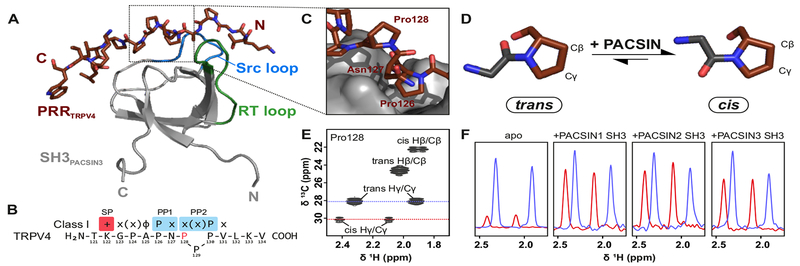
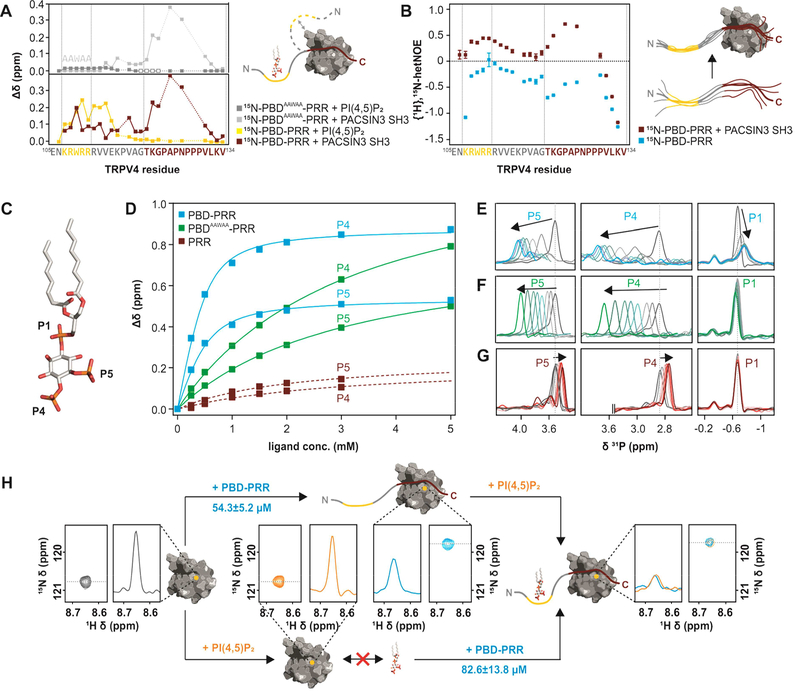
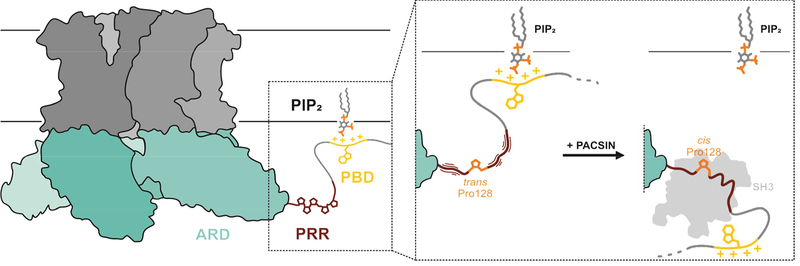
Transient receptor potential (TRP) channels are polymodally regulated ion channels. TRPV4 (vanilloid 4) is sensitized by PIP2 and desensitized by Syndapin3/PACSIN3, which bind to the structurally uncharacterized TRPV4 N terminus. We determined the nuclear magnetic resonance structure of the Syndapin3/PACSIN3 SH3 domain in complex with the TRPV4 N-terminal proline-rich region (PRR), which binds as a class I polyproline II (PPII) helix. This PPII conformation is broken by a conserved proline in a cis conformation. Beyond the PPII, we find that the proximal TRPV4 N terminus is unstructured, a feature conserved across species thus explaining the difficulties in resolving it in previous structural studies. Syndapin/PACSIN SH3 domain binding leads to rigidification of both the PRR and the adjacent PIP2 binding site. We determined the affinities of the TRPV4 N terminus for PACSIN1, 2, and 3 SH3 domains and PIP2 and deduce a hierarchical interaction network where Syndapin/PACSIN binding influences the PIP2 binding site but not vice versa.
Keywords: NMR; PACSIN; PIP(2); SH3 domain; Syndapin; TRP channel; TRPV4; cis proline; class I; proline-rich region.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Figures
References
-
- Afonin S, Kubyshkin V, Mykhailiuk PK, Komarov IV, and Ulrich AS (2017). Conformational Plasticity of the Cell-Penetrating Peptide SAP As Revealed by Solid-State19F-NMR and Circular Dichroism Spectroscopies. The journal of physical chemistry. B 121, 6479–6491. - PubMed
-
- Bhattacharya A, Tejero R, and Montelione GT (2007). Evaluating protein structures determined by structural genomics consortia. Proteins 66, 778–795. - PubMed
-
- Bokhovchuk FM, Bate N, Kovalevskaya NV, Goult BT, Spronk CAEM, and Vuister GW (2018). The Structural Basis of Calcium Dependent Inactivation of the Transient Receptor Potential Vanilloid 5 Channel. Biochemistry. - PubMed
-
- Breeze AL (2000). Isotope-filtered NMR methods for the study of biomolecular structure and interactions. Progress in nuclear magnetic resonance spectroscopy 36, 323–372.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
