

A Neural Circuit for Gut-Induced Reward
- PMID: 30245012
- PMCID: PMC6195474
- DOI: 10.1016/j.cell.2018.08.049
A Neural Circuit for Gut-Induced Reward
Erratum in
-
A Neural Circuit for Gut-Induced Reward.Cell. 2018 Oct 18;175(3):887-888. doi: 10.1016/j.cell.2018.10.018. Cell. 2018. PMID: 30340046 No abstract available.
Abstract
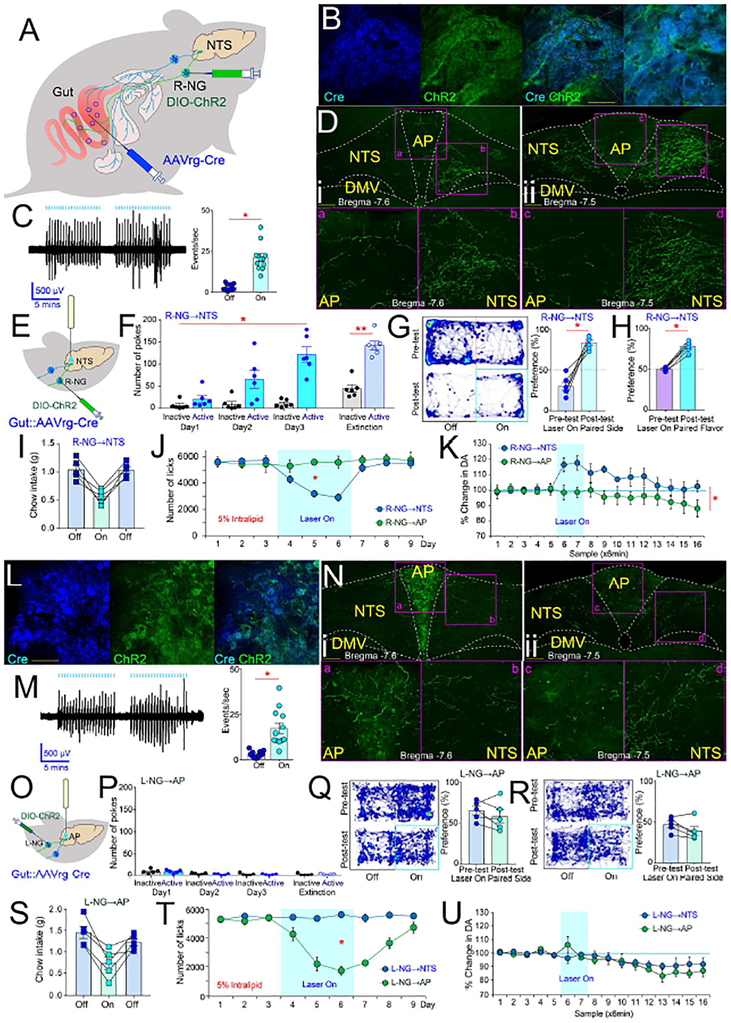
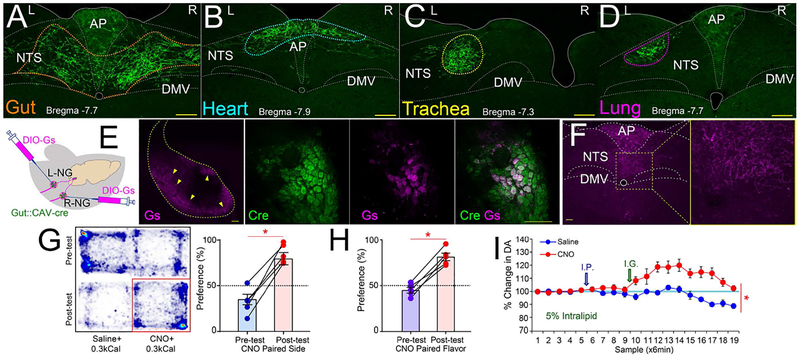
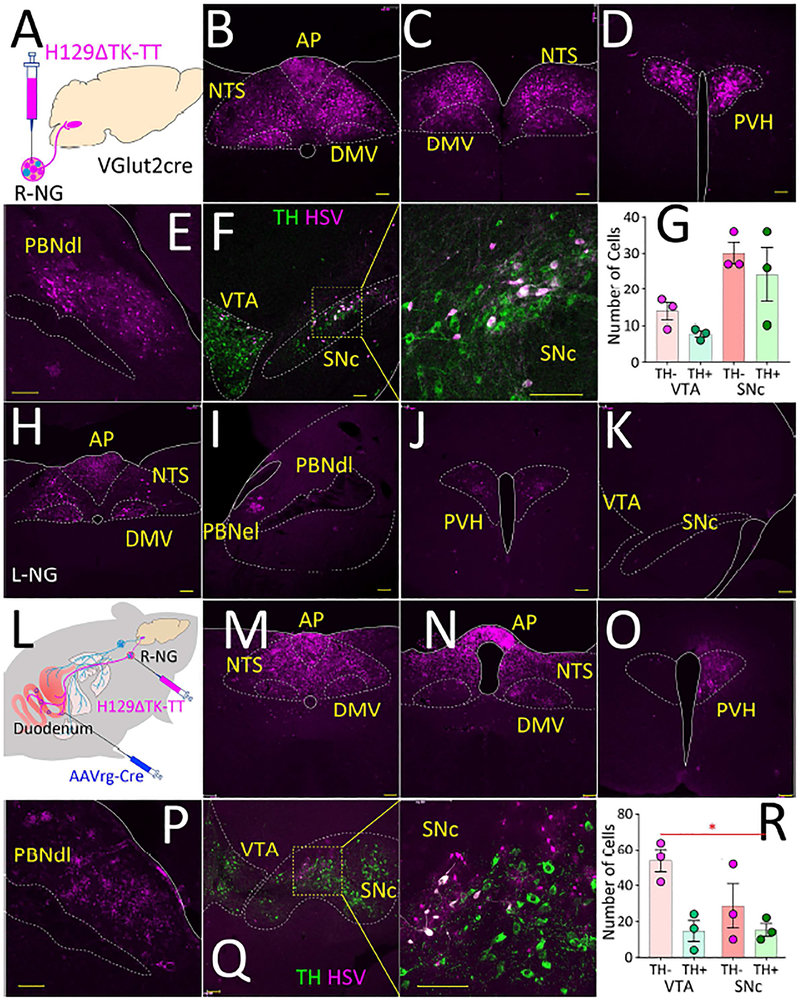
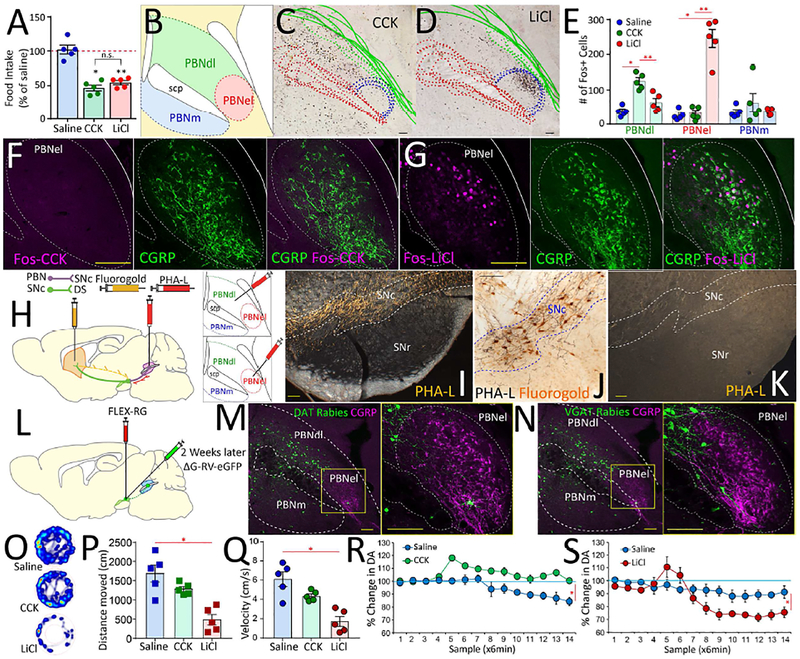
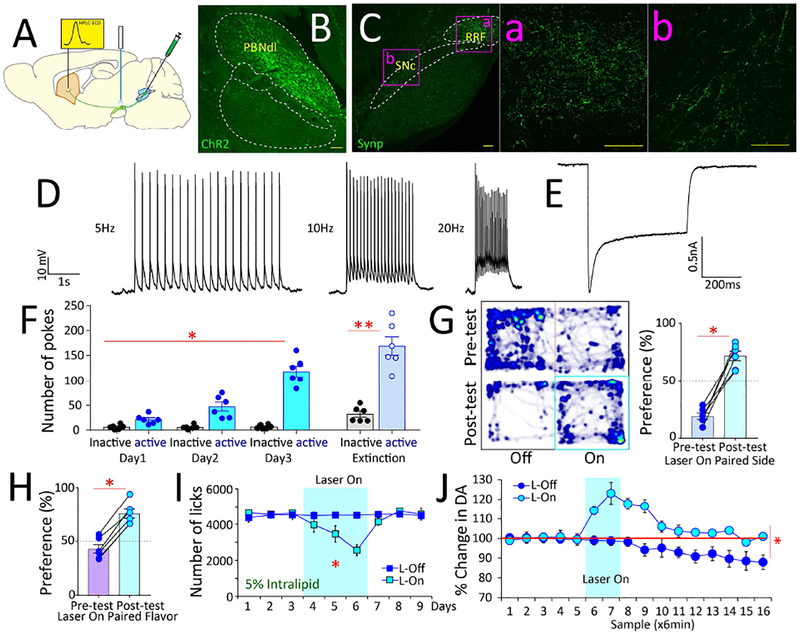
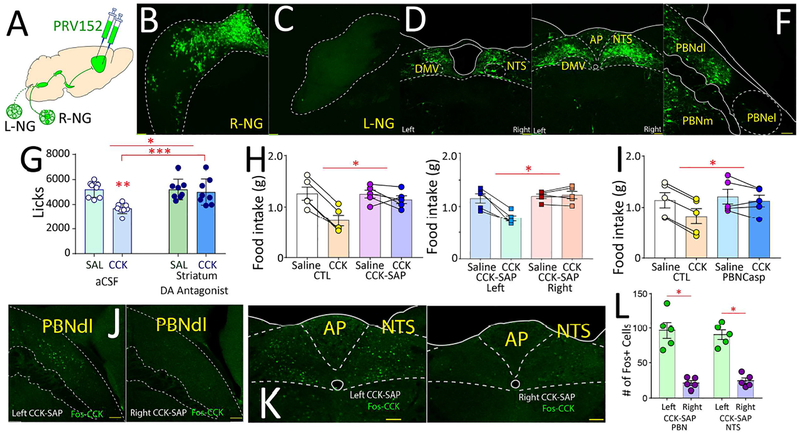
The gut is now recognized as a major regulator of motivational and emotional states. However, the relevant gut-brain neuronal circuitry remains unknown. We show that optical activation of gut-innervating vagal sensory neurons recapitulates the hallmark effects of stimulating brain reward neurons. Specifically, right, but not left, vagal sensory ganglion activation sustained self-stimulation behavior, conditioned both flavor and place preferences, and induced dopamine release from Substantia nigra. Cell-specific transneuronal tracing revealed asymmetric ascending pathways of vagal origin throughout the CNS. In particular, transneuronal labeling identified the glutamatergic neurons of the dorsolateral parabrachial region as the obligatory relay linking the right vagal sensory ganglion to dopamine cells in Substantia nigra. Consistently, optical activation of parabrachio-nigral projections replicated the rewarding effects of right vagus excitation. Our findings establish the vagal gut-to-brain axis as an integral component of the neuronal reward pathway. They also suggest novel vagal stimulation approaches to affective disorders.
Keywords: dopamine; gut-brain axis; reward; vagus nerve.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures
Comment in
-
Rewarding gut feeling.Nat Rev Neurosci. 2018 Nov;19(11):639. doi: 10.1038/s41583-018-0075-3. Nat Rev Neurosci. 2018. PMID: 30291300 No abstract available.
References
-
- Altschuler SM, Bao XM, Bieger D, Hopkins DA, and Miselis RR (1989). Viscerotopic representation of the upper alimentary tract in the rat: sensory ganglia and nuclei of the solitary and spinal trigeminal tracts. J Comp Neur 283, 248–268. - PubMed
-
- Angyan L (1975). Vagal influences on hypothalamic self-stimulation in the cat. Life Sci 17, 289–292. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
