

Bi-allelic Mutations in NDUFA6 Establish Its Role in Early-Onset Isolated Mitochondrial Complex I Deficiency
- PMID: 30245030
- PMCID: PMC6174280
- DOI: 10.1016/j.ajhg.2018.08.013
Bi-allelic Mutations in NDUFA6 Establish Its Role in Early-Onset Isolated Mitochondrial Complex I Deficiency
Abstract
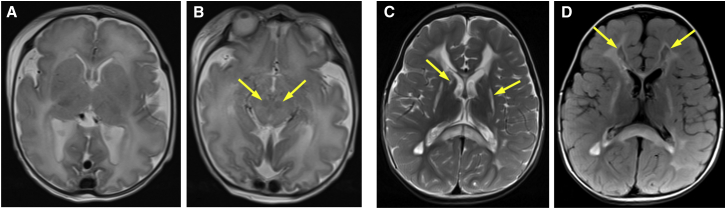
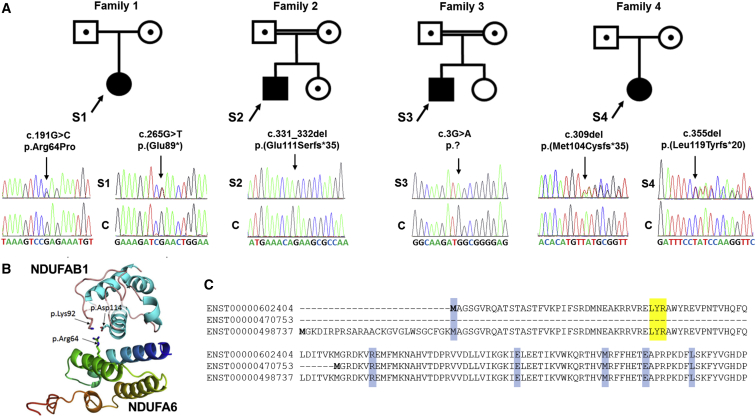
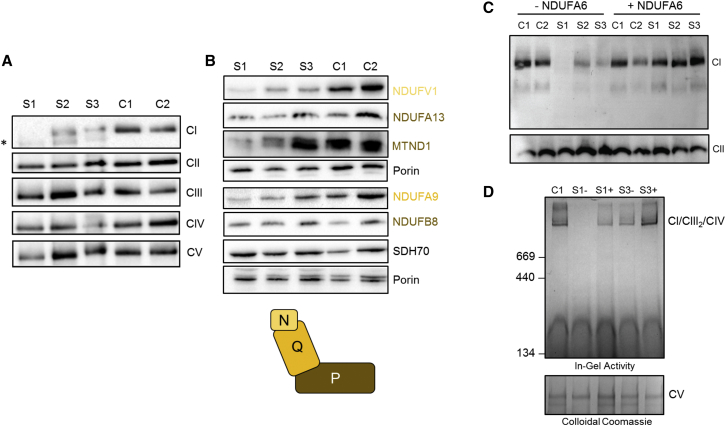
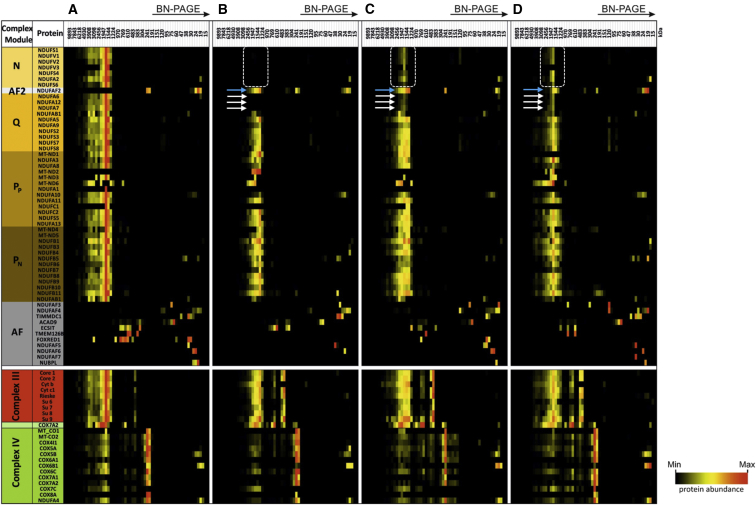
Isolated complex I deficiency is a common biochemical phenotype observed in pediatric mitochondrial disease and often arises as a consequence of pathogenic variants affecting one of the ∼65 genes encoding the complex I structural subunits or assembly factors. Such genetic heterogeneity means that application of next-generation sequencing technologies to undiagnosed cohorts has been a catalyst for genetic diagnosis and gene-disease associations. We describe the clinical and molecular genetic investigations of four unrelated children who presented with neuroradiological findings and/or elevated lactate levels, highly suggestive of an underlying mitochondrial diagnosis. Next-generation sequencing identified bi-allelic variants in NDUFA6, encoding a 15 kDa LYR-motif-containing complex I subunit that forms part of the Q-module. Functional investigations using subjects' fibroblast cell lines demonstrated complex I assembly defects, which were characterized in detail by mass-spectrometry-based complexome profiling. This confirmed a marked reduction in incorporated NDUFA6 and a concomitant reduction in other Q-module subunits, including NDUFAB1, NDUFA7, and NDUFA12. Lentiviral transduction of subjects' fibroblasts showed normalization of complex I. These data also support supercomplex formation, whereby the ∼830 kDa complex I intermediate (consisting of the P- and Q-modules) is in complex with assembled complex III and IV holoenzymes despite lacking the N-module. Interestingly, RNA-sequencing data provided evidence that the consensus RefSeq accession number does not correspond to the predominant transcript in clinically relevant tissues, prompting revision of the NDUFA6 RefSeq transcript and highlighting not only the importance of thorough variant interpretation but also the assessment of appropriate transcripts for analysis.
Keywords: NDUFA6; complex I; complexome profiling; mitochondrial disease.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Formosa L.E., Dibley M.G., Stroud D.A., Ryan M.T. Building a complex complex: Assembly of mitochondrial respiratory chain complex I. Semin. Cell Dev. Biol. 2018;76:154–162. - PubMed
-
- Piekutowska-Abramczuk D., Assouline Z., Mataković L., Feichtinger R.G., Koňařiková E., Jurkiewicz E., Stawiński P., Gusic M., Koller A., Pollak A. NDUFB8 mutations cause mitochondrial complex I deficiency in individuals with Leigh-like encephalomyopathy. Am. J. Hum. Genet. 2018;102:460–467. - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
