

Sources of all-trans retinal oxidation independent of the aldehyde dehydrogenase 1A isozymes exist in the postnatal testis†
- PMID: 30247516
- PMCID: PMC6378859
- DOI: 10.1093/biolre/ioy200
Sources of all-trans retinal oxidation independent of the aldehyde dehydrogenase 1A isozymes exist in the postnatal testis†
Abstract
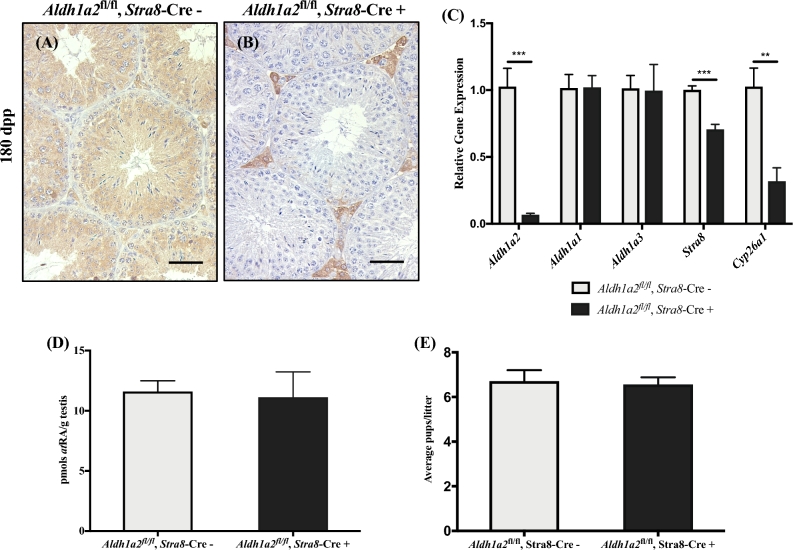
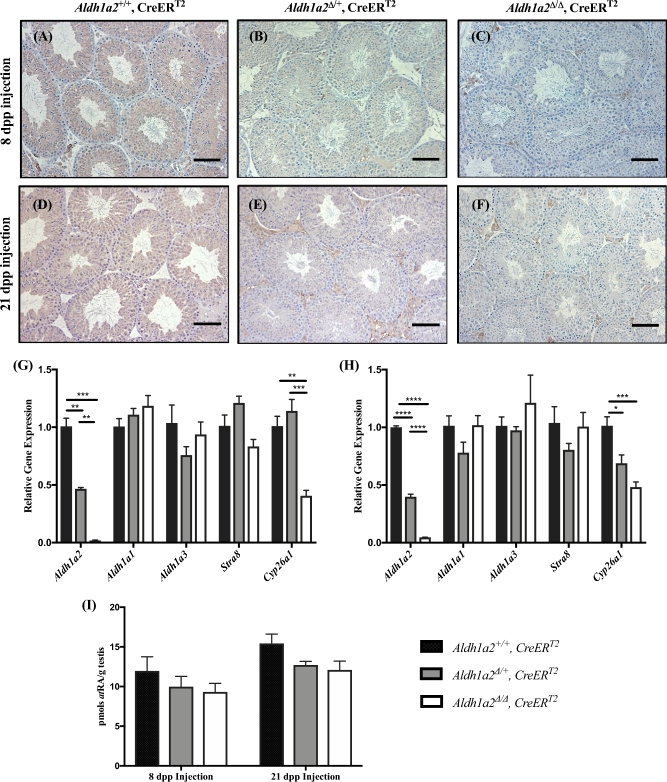
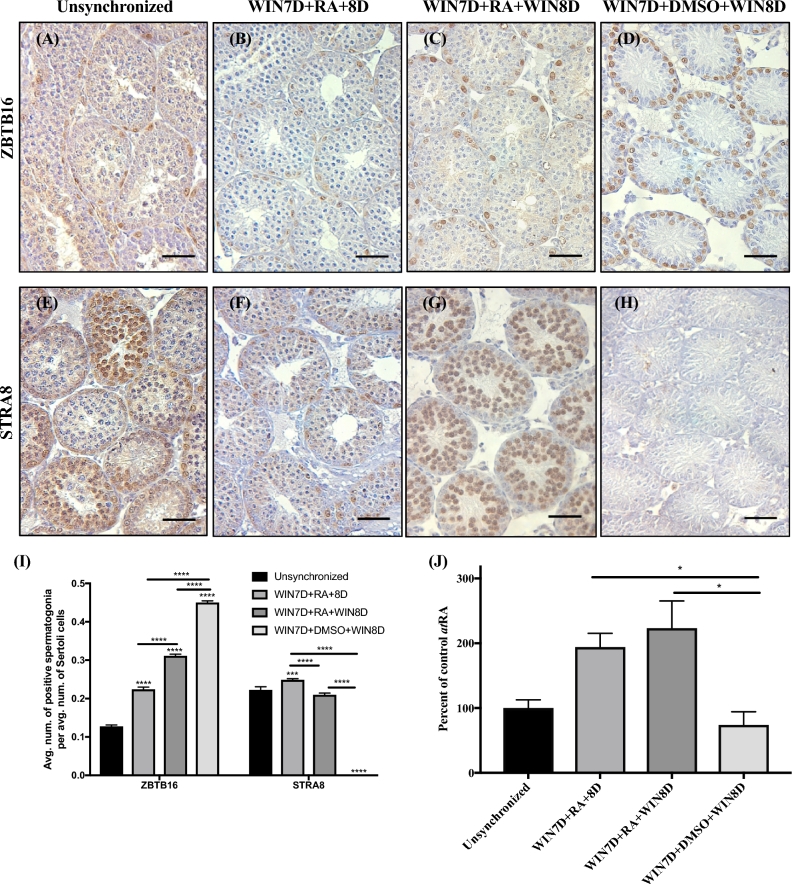
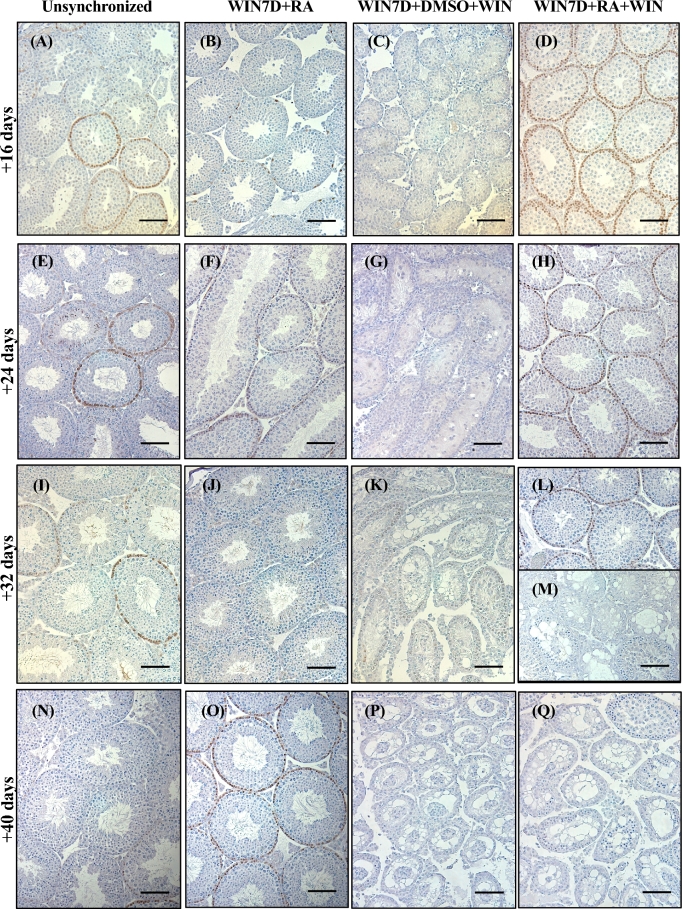
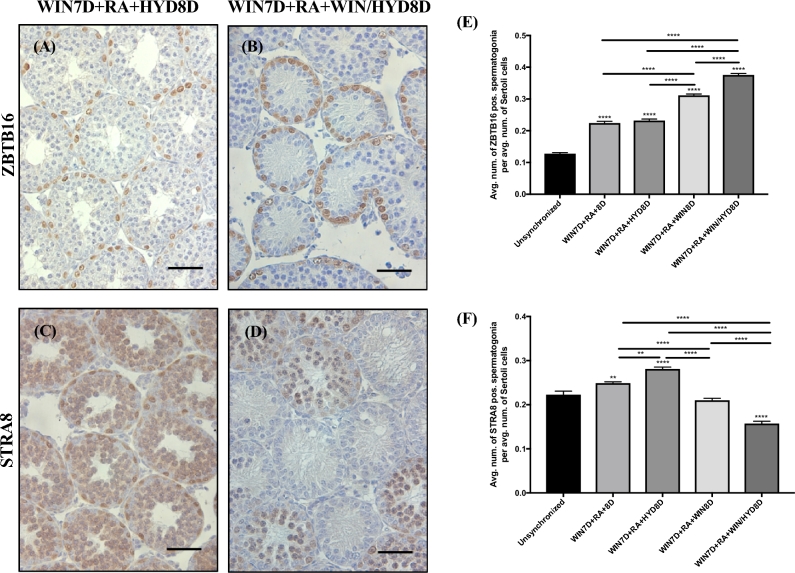
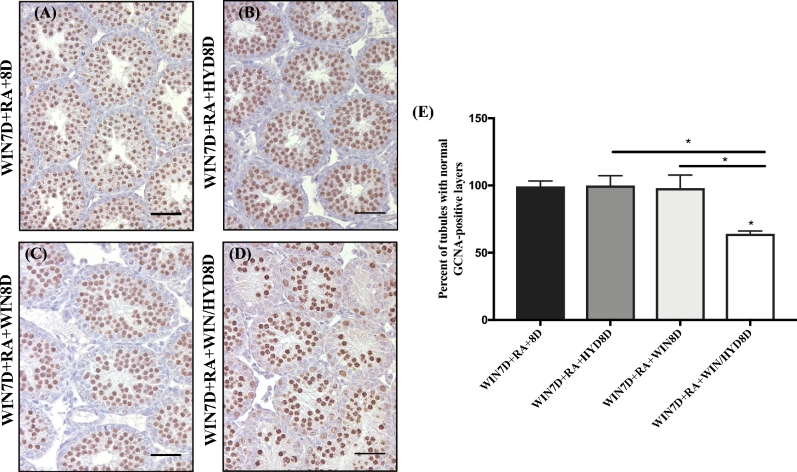
Despite the essential role of the active metabolite of vitamin A, all-trans retinoic acid (atRA) in spermatogenesis, the enzymes, and cellular populations responsible for its synthesis in the postnatal testis remain largely unknown. The aldehyde dehydrogenase 1A (ALDH1A) family of enzymes residing within Sertoli cells is responsible for the synthesis of atRA, driving the first round of spermatogenesis. Those studies also revealed that the atRA required to drive subsequent rounds of spermatogenesis is possibly derived from the ALDH1A enzymes residing within the meiotic and post-meiotic germ cells. Three ALDH1A isozymes (ALDH1A1, ALDH1A2, and ALDH1A3) are present in the testis. Although, ALDH1A1 is expressed in adult Sertoli cells and is suggested to contribute to the atRA required for the pre-meiotic transitions, ALDH1A2 is proposed to be the essential isomer involved in testicular atRA biosynthesis. In this report, we first examine the requirement for ALDH1A2 via the generation and analysis of a conditional Aldh1a2 germ cell knockout and a tamoxifen-induced Aldh1a2 knockout model. We then utilized the pan-ALDH1A inhibitor (WIN 18446) to test the collective contribution of the ALDH1A enzymes to atRA biosynthesis following the first round of spermatogenesis. Collectively, our data provide the first in vivo evidence demonstrating that animals severely deficient in ALDH1A2 postnatally proceed normally through spermatogenesis. Our studies with a pan-ALDH1A inhibitor (WIN 18446) also suggest that an alternative source of atRA biosynthesis independent of the ALDH1A enzymes becomes available to maintain atRA levels for several spermatogenic cycles following an initial atRA injection.
Keywords: Aldh1A; Aldh1a2; WIN 18,446; retinoic acid; spermatogenesis; spermatogonia; testis.
© The Author(s) 2018. Published by Oxford University Press on behalf of Society for the Study of Reproduction.
Figures
References
-
- Yang QE, Oatley JM. Spermatogonial stem cell functions in physiological and pathological conditions. Curr Top Dev Biol 2014; 107:235–267. - PubMed
-
- Drumond AL, Meistrich ML, Chiarini-Garcia H. Spermatogonial morphology and kinetics during testis development in mice: a high-resolution light microscopy approach. Reproduction 2011; 142:145–155. - PubMed
-
- Oakberg EF. A description of spermiogenesis in the mouse and its use in analysis of the cycle of the seminiferous epithelium and germ cell renewal. Am J Anat 1956; 99:391–413. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
