

Key residues controlling bidirectional ion movements in Na+/Ca2+ exchanger
- PMID: 30248574
- PMCID: PMC6688843
- DOI: 10.1016/j.ceca.2018.09.004
Key residues controlling bidirectional ion movements in Na+/Ca2+ exchanger
Abstract
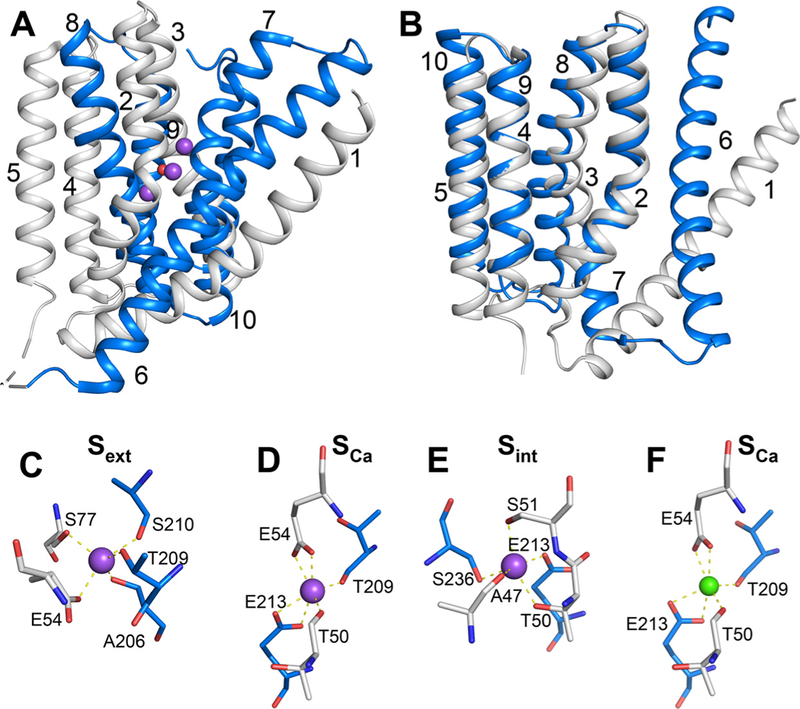
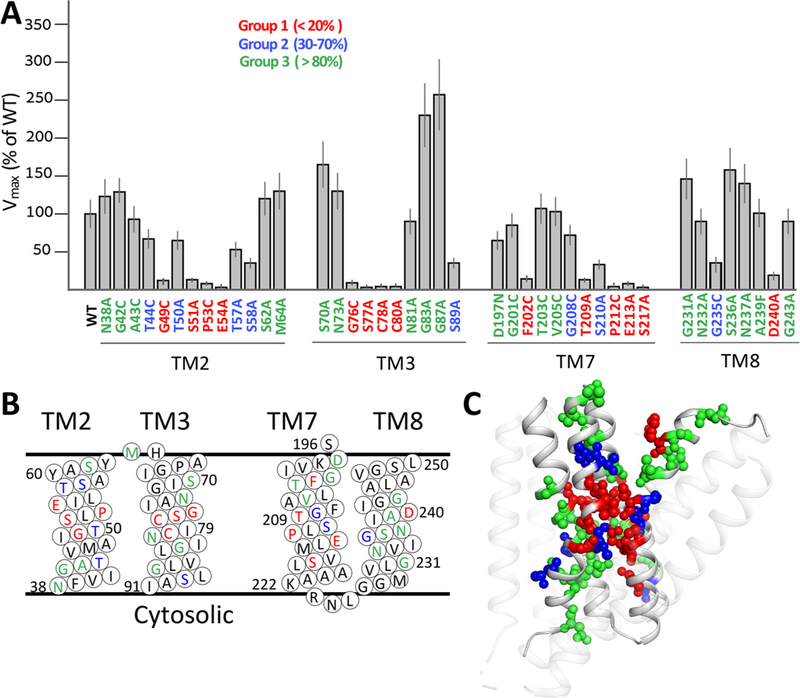
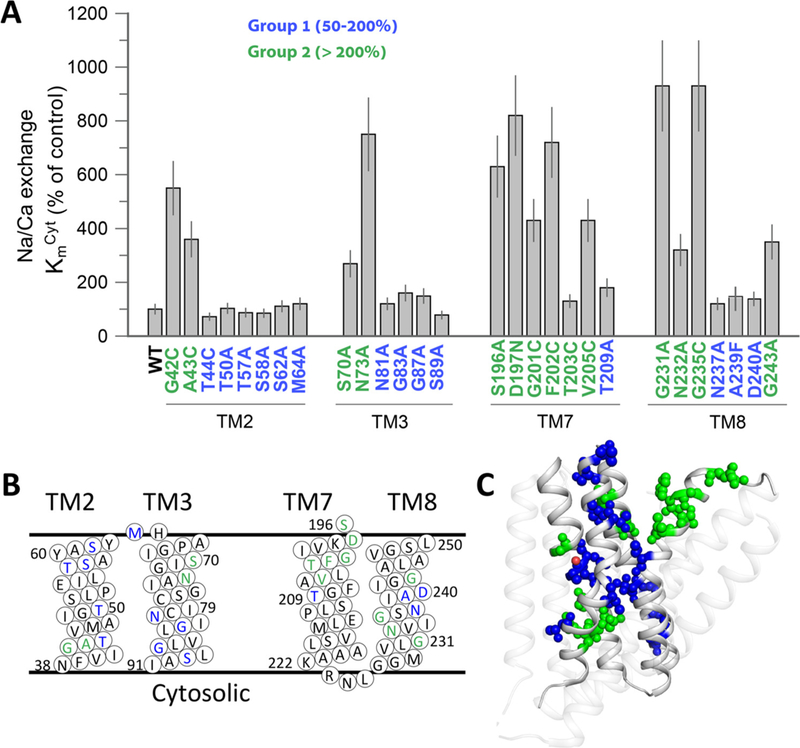
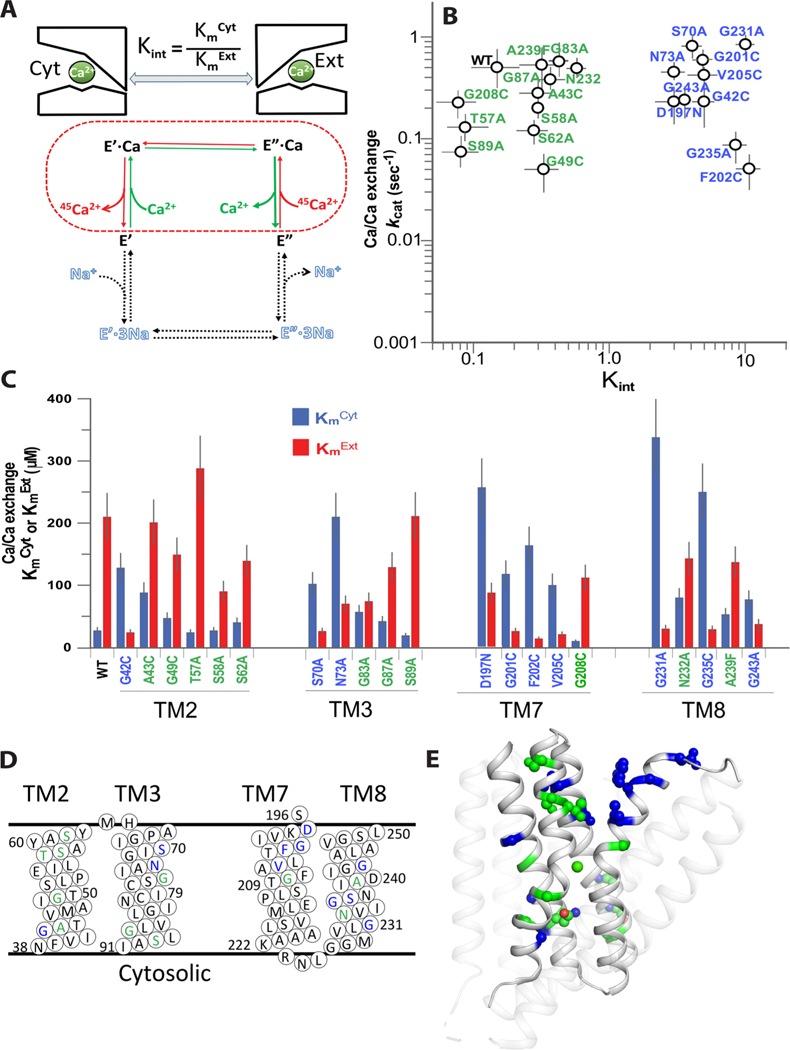
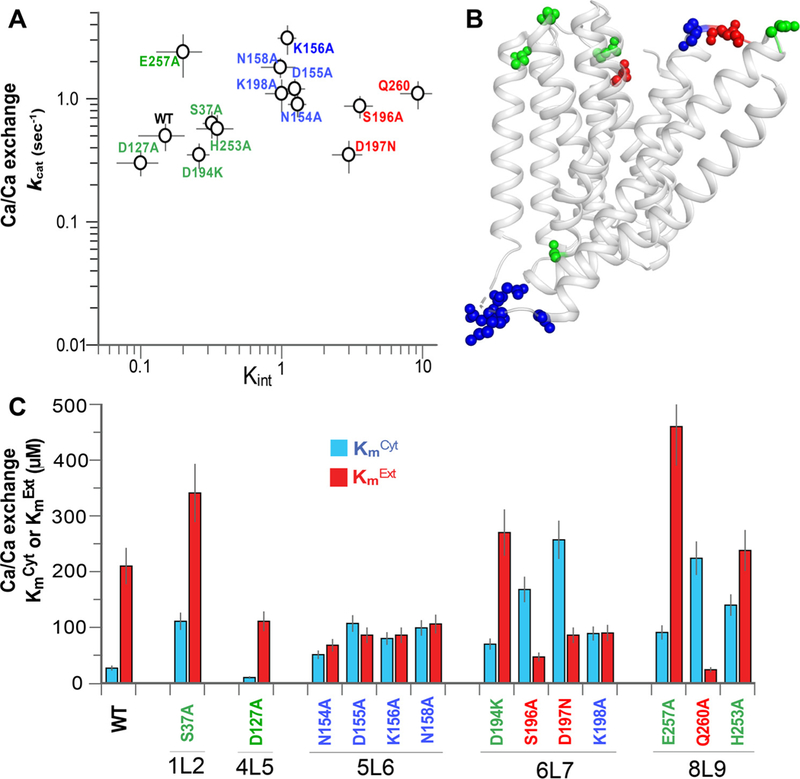
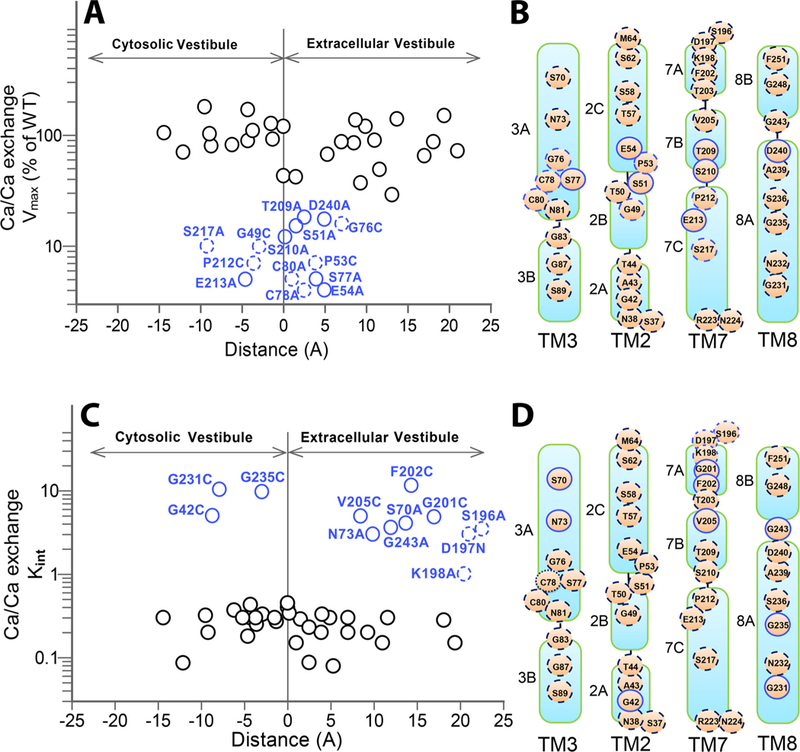
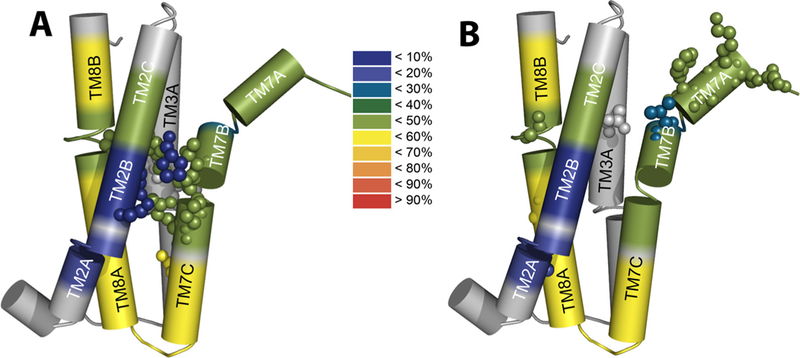
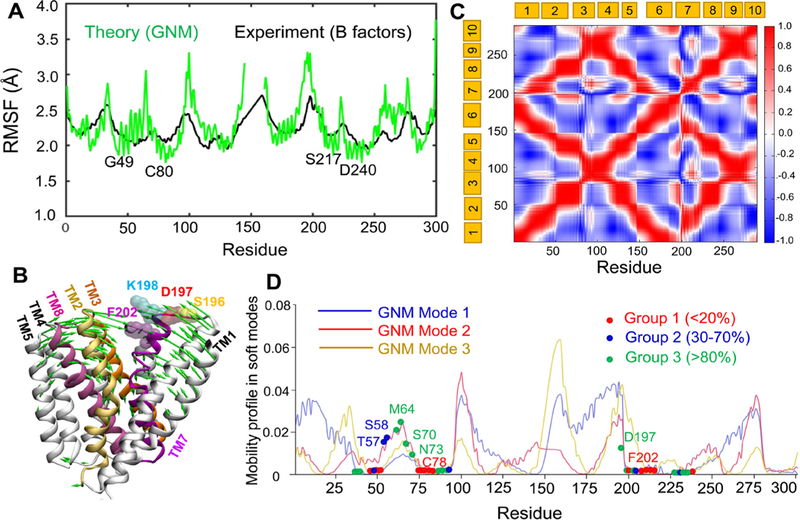
Prokaryotic and eukaryotic Na+/Ca2+ exchangers (NCX) control Ca2+ homeostasis. NCX orthologs exhibit up to 104-fold differences in their turnover rates (kcat), whereas the ratios between the cytosolic (cyt) and extracellular (ext) Km values (Kint = KmCyt/KmExt) are highly asymmetric and alike (Kint ≤ 0.1) among NCXs. The structural determinants controlling a huge divergence in kcat at comparable Kint remain unclear, although 11 (out of 12) ion-coordinating residues are highly conserved among NCXs. The crystal structure of the archaeal NCX (NCX_Mj) was explored for testing the mutational effects of pore-allied and loop residues on kcat and Kint. Among 55 tested residues, 26 mutations affect either kcat or Kint, where two major groups can be distinguished. The first group of mutations (14 residues) affect kcat rather than Kint. The majority of these residues (10 out of 14) are located within the extracellular vestibule near the pore center. The second group of mutations (12 residues) affect Kint rather than kcat, whereas the majority of residues (9 out 12) are randomly dispersed within the extracellular vestibule. In conjunction with computational modeling-simulations and hydrogen-deuterium exchange mass-spectrometry (HDX-MS), the present mutational analysis highlights structural elements that differentially govern the intrinsic asymmetry and transport rates. The key residues, located at specific segments, can affect the characteristic features of local backbone dynamics and thus, the conformational flexibility of ion-transporting helices contributing to critical conformational transitions. The underlying mechanisms might have a physiological relevance for matching the response modes of NCX variants to cell-specific Ca2+ and Na+ signaling.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Conflict of interest
All authors state no conflict of interests.
Figures
References
-
- Philipson KD, Nicoll DA, Sodium-calcium exchange: a molecular perspective, Annu. Rev. Physiol. 62 (2000) 111–133. - PubMed
-
- Blaustein MP, Lederer WJ, Sodium/calcium exchange: its physiological implications, Physiol. Rev. 79 (1999) 763–854. - PubMed
-
- Khananshvili D, The SLC8 gene family of sodium-calcium exchangers (NCX): structure, function, and regulation in health and disease, Mol. Asp. Med. 34 (2013) 220–235. - PubMed
-
- Palty R, Silverman WF, Hershfinkel M, Caporale T, Sensi SL, Parnis J, Nolte C, Fishman D, Shoshan-Barmatz V, Herrmann S, Khananshvili D, Sekler I, NCLX is an essential component of mitochondrial Na+/Ca2+ exchange, Proc. Natl. Acad. Sci. U. S. A. 107 (2010) 436–441, 10.1073/pnas.0908099107. - DOI - PMC - PubMed
-
- Khananshvili D, Sodium-calcium exchangers (NCX): molecular hallmarks underlying the tissue-specific and systemic functions, Plügers Arch. 466 (2014) 43–60. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
