

Identity and novelty in the avian syrinx
- PMID: 30249637
- PMCID: PMC6187200
- DOI: 10.1073/pnas.1804586115
Identity and novelty in the avian syrinx
Abstract
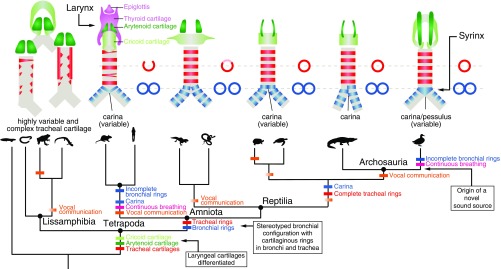
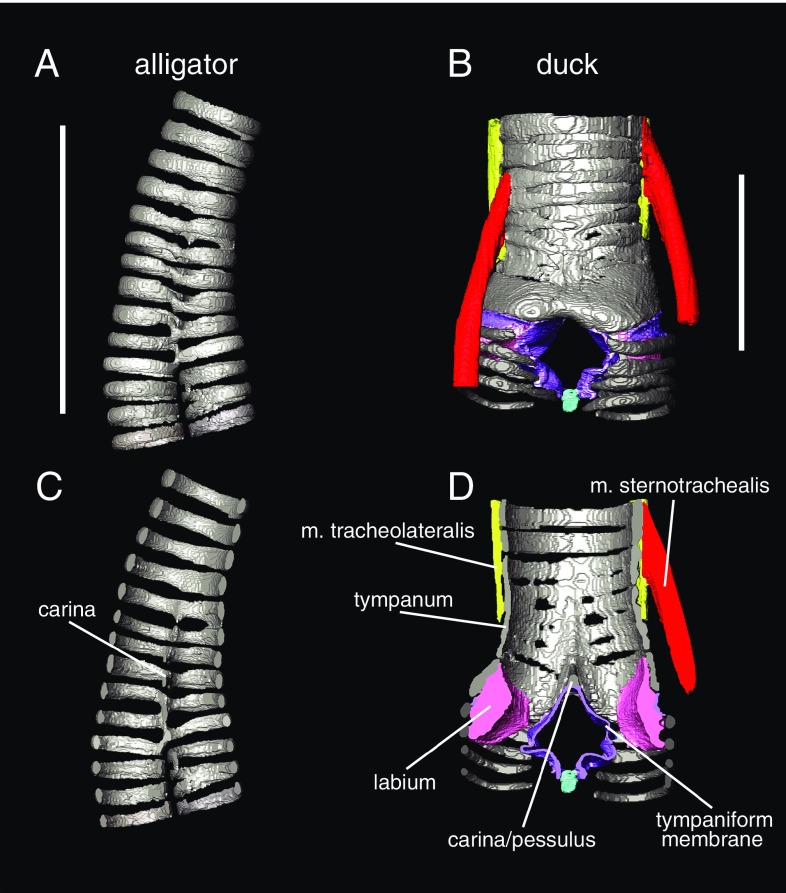
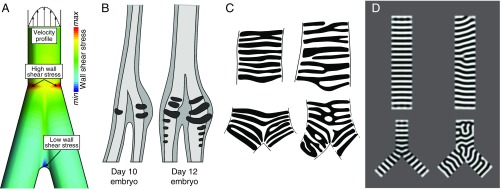
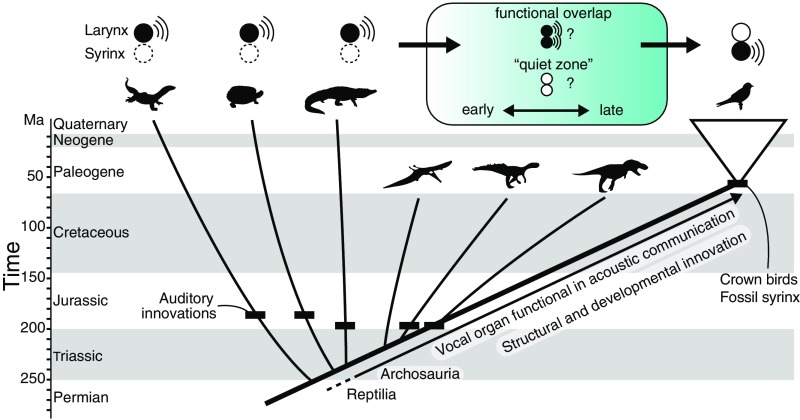
In its most basic conception, a novelty is simply something new. However, when many previously proposed evolutionary novelties have been illuminated by genetic, developmental, and fossil data, they have refined and narrowed our concept of biological "newness." For example, they show that these novelties can occur at one or multiple levels of biological organization. Here, we review the identity of structures in the avian vocal organ, the syrinx, and bring together developmental data on airway patterning, structural data from across tetrapods, and mathematical modeling to assess what is novel. In contrast with laryngeal cartilages that support vocal folds in other vertebrates, we find no evidence that individual cartilage rings anchoring vocal folds in the syrinx have homology with any specific elements in outgroups. Further, unlike all other vertebrate vocal organs, the syrinx is not derived from a known valve precursor, and its origin involves a transition from an evolutionary "spandrel" in the respiratory tract, the site where the trachea meets the bronchi, to a target for novel selective regimes. We find that the syrinx falls into an unusual category of novel structures: those having significant functional overlap with the structures they replace. The syrinx, along with other evolutionary novelties in sensory and signaling modalities, may more commonly involve structural changes that contribute to or modify an existing function rather than those that enable new functions.
Keywords: bioacoustics; birds; tetrapods; tracheal rings; vocal communication.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Xu X, et al. An integrative approach to understanding bird origins. Science. 2014;346:1253293. - PubMed
-
- Muller GB, Wagner GP. Novelty in evolution: Restructuring the concept. Annu Rev Ecol Syst. 1991;22:229–256.
-
- Shubin N, Tabin C, Carroll S. Fossils, genes and the evolution of animal limbs. Nature. 1997;388:639–648. - PubMed
-
- Hall BK, Kerney R. Levels of biological organization and the origin of novelty. J Exp Zoolog B Mol Dev Evol. 2012;318:428–437. - PubMed
-
- Riede T, Li Z, Tokuda IT, Farmer CG. Functional morphology of the Alligator mississippiensis larynx with implications for vocal production. J Exp Biol. 2015;218:991–998. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
