

Human Endogenous Retrovirus-K HML-2 integration within RASGRF2 is associated with intravenous drug abuse and modulates transcription in a cell-line model
- PMID: 30249655
- PMCID: PMC6187174
- DOI: 10.1073/pnas.1811940115
Human Endogenous Retrovirus-K HML-2 integration within RASGRF2 is associated with intravenous drug abuse and modulates transcription in a cell-line model
Abstract
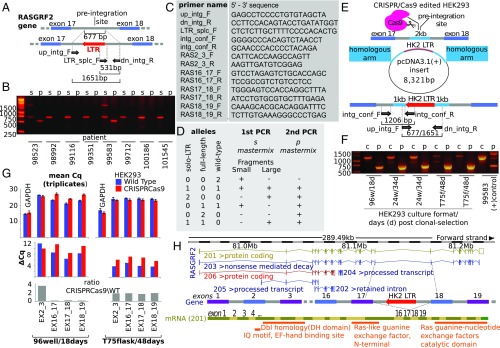
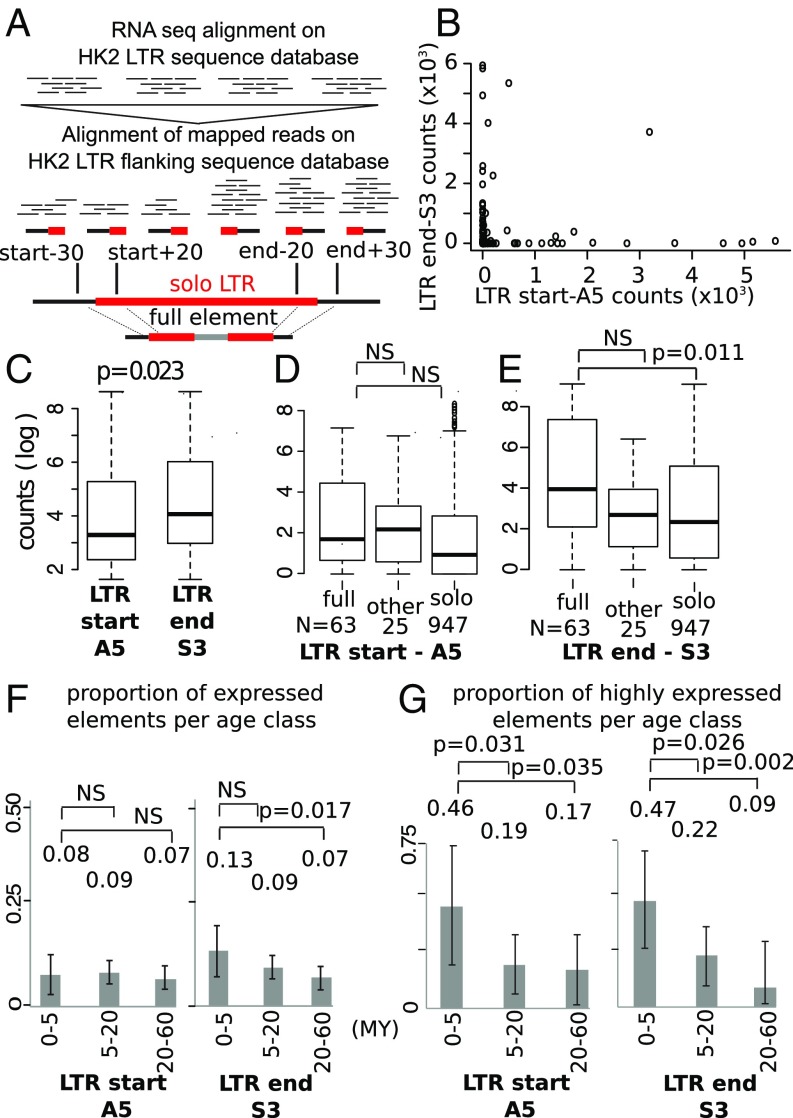
HERV-K HML-2 (HK2) has been proliferating in the germ line of humans at least as recently as 250,000 years ago, with some integrations that remain polymorphic in the modern human population. One of the solitary HK2 LTR polymorphic integrations lies between exons 17 and 18 of RASGRF2, a gene that affects dopaminergic activity and is thus related to addiction. Here we show that this antisense HK2 integration (namely RASGRF2-int) is found more frequently in persons who inject drugs compared with the general population. In a Greek HIV-1-positive population (n = 202), we found RASGRF2-int 2.5 times (14 versus 6%) more frequently in patients infected through i.v. drug use compared with other transmission route controls (P = 0.03). Independently, in a United Kingdom-based hepatitis C virus-positive population (n = 184), we found RASGRF2-int 3.6 times (34 versus 9.5%) more frequently in patients infected during chronic drug abuse compared with controls (P < 0.001). We then tested whether RASGRF2-int could be mechanistically responsible for this association by modulating transcription of RASGRF2 We show that the CRISPR/Cas9-mediated insertion of HK2 in HEK293 cells in the exact RASGRF2 intronic position found in the population resulted in significant transcriptional and phenotypic changes. We also explored mechanistic features of other intronic HK2 integrations and show that HK2 LTRs can be responsible for generation of cis-natural antisense transcripts, which could interfere with the transcription of nearby genes. Our findings suggest that RASGRF2-int is a strong candidate for dopaminergic manipulation, and emphasize the importance of accurate mapping of neglected HERV polymorphisms in human genomic studies.
Keywords: HERV-K HML-2; RASGRF2; addiction; endogenous retrovirus; persons who inject drugs.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Human Endogenous Retrovirus K (HML-2) in Health and Disease.Front Microbiol. 2020 Jul 17;11:1690. doi: 10.3389/fmicb.2020.01690. eCollection 2020. Front Microbiol. 2020. PMID: 32765477 Free PMC article. Review.
-
Human Endogenous Retrovirus Type K (HERV-K) Particles Package and Transmit HERV-K-Related Sequences.J Virol. 2015 Jul;89(14):7187-201. doi: 10.1128/JVI.00544-15. Epub 2015 Apr 29. J Virol. 2015. PMID: 25926654 Free PMC article.
-
CpG methylation directly regulates transcriptional activity of the human endogenous retrovirus family HERV-K(HML-2).J Virol. 2005 Jan;79(2):876-83. doi: 10.1128/JVI.79.2.876-883.2005. J Virol. 2005. PMID: 15613316 Free PMC article.
-
Mechanisms of HERV-K (HML-2) Transcription during Human Mammary Epithelial Cell Transformation.J Virol. 2017 Dec 14;92(1):e01258-17. doi: 10.1128/JVI.01258-17. Print 2018 Jan 1. J Virol. 2017. PMID: 29046454 Free PMC article.
-
Human Endogenous Retrovirus-K (HML-2)-Related Genetic Variation: Human Genome Diversity and Disease.Genes (Basel). 2023 Nov 28;14(12):2150. doi: 10.3390/genes14122150. Genes (Basel). 2023. PMID: 38136972 Free PMC article. Review.
Cited by
-
Genotyping of Transposable Element Insertions Segregating in Human Populations Using Short-Read Realignments.Methods Mol Biol. 2023;2607:63-83. doi: 10.1007/978-1-0716-2883-6_4. Methods Mol Biol. 2023. PMID: 36449158
-
Not all viruses cause disease: HERV-K(HML-2) in healthy human tissues.PLoS Biol. 2022 Oct 31;20(10):e3001884. doi: 10.1371/journal.pbio.3001884. eCollection 2022 Oct. PLoS Biol. 2022. PMID: 36315578 Free PMC article.
-
Human Endogenous Retrovirus K (HML-2) in Health and Disease.Front Microbiol. 2020 Jul 17;11:1690. doi: 10.3389/fmicb.2020.01690. eCollection 2020. Front Microbiol. 2020. PMID: 32765477 Free PMC article. Review.
-
Implications of Endogenous Retroelements in the Etiopathogenesis of Systemic Lupus Erythematosus.J Clin Med. 2021 Feb 19;10(4):856. doi: 10.3390/jcm10040856. J Clin Med. 2021. PMID: 33669709 Free PMC article. Review.
-
Characterization of the brain virome in human immunodeficiency virus infection and substance use disorder.PLoS One. 2024 Apr 17;19(4):e0299891. doi: 10.1371/journal.pone.0299891. eCollection 2024. PLoS One. 2024. PMID: 38630782 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
