

Regulation of the Natriuretic Peptide Receptor 2 (Npr2) by Phosphorylation of Juxtamembrane Serine and Threonine Residues Is Essential for Bifurcation of Sensory Axons
- PMID: 30249793
- PMCID: PMC6222061
- DOI: 10.1523/JNEUROSCI.0495-18.2018
Regulation of the Natriuretic Peptide Receptor 2 (Npr2) by Phosphorylation of Juxtamembrane Serine and Threonine Residues Is Essential for Bifurcation of Sensory Axons
Abstract
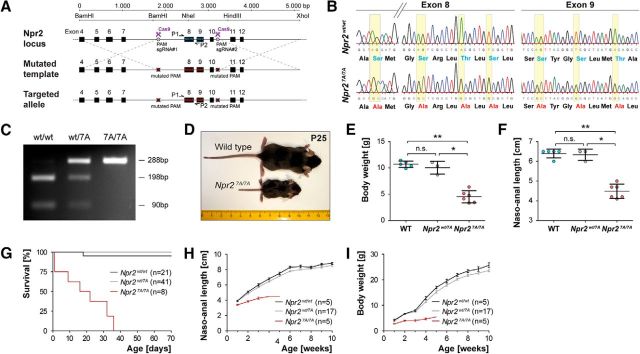
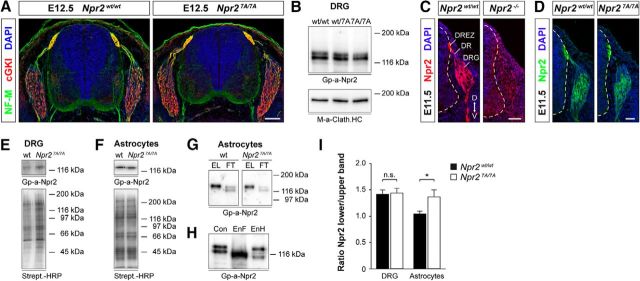
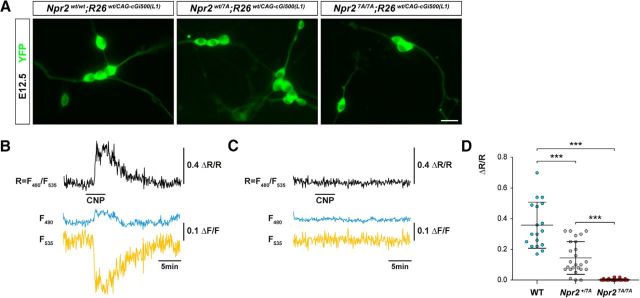
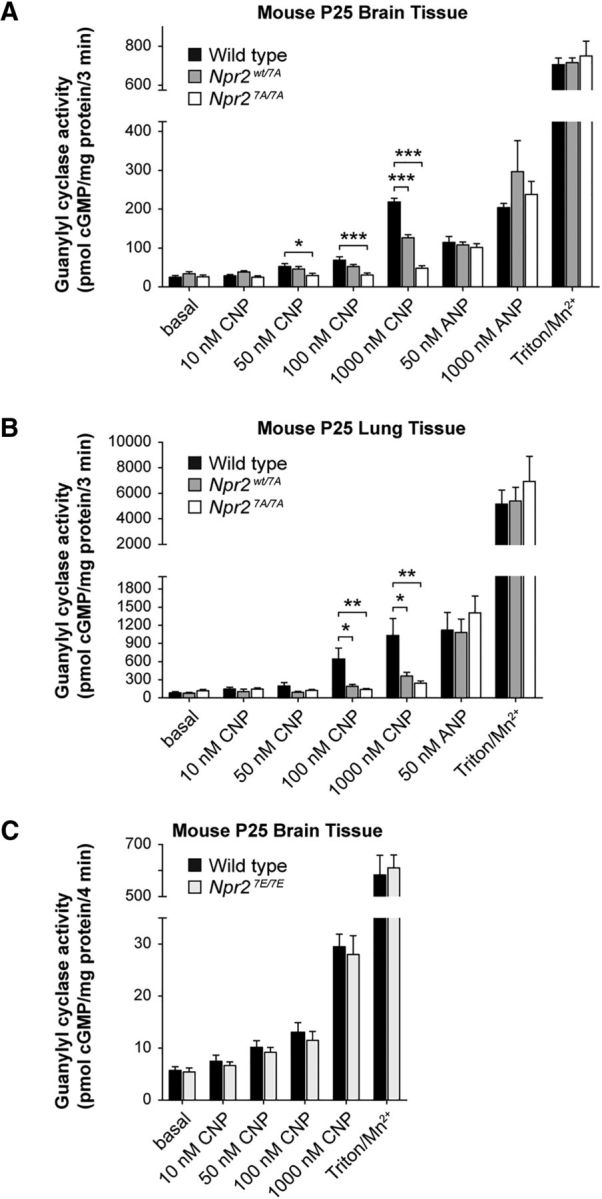
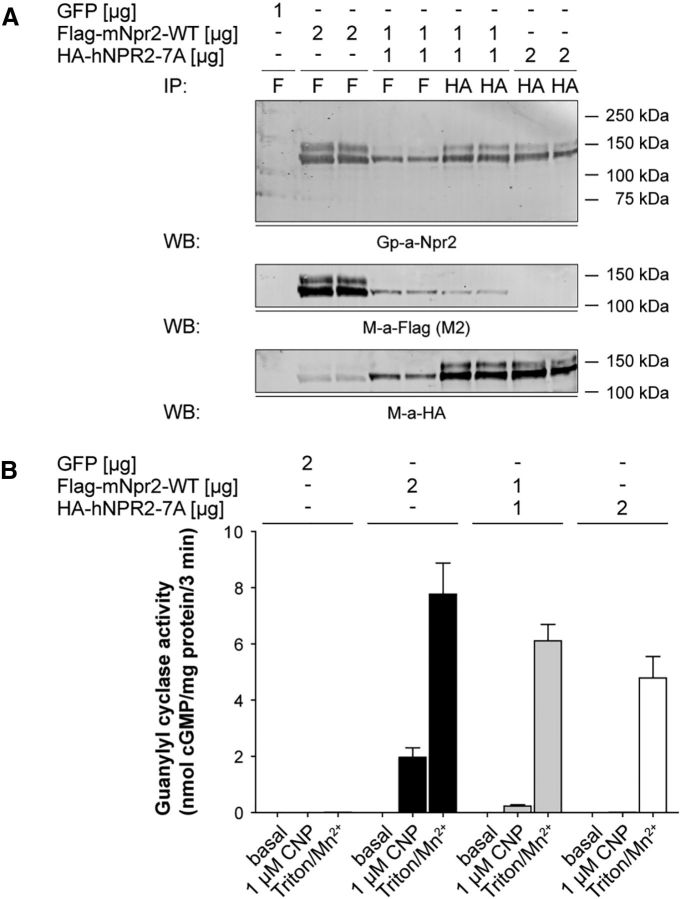
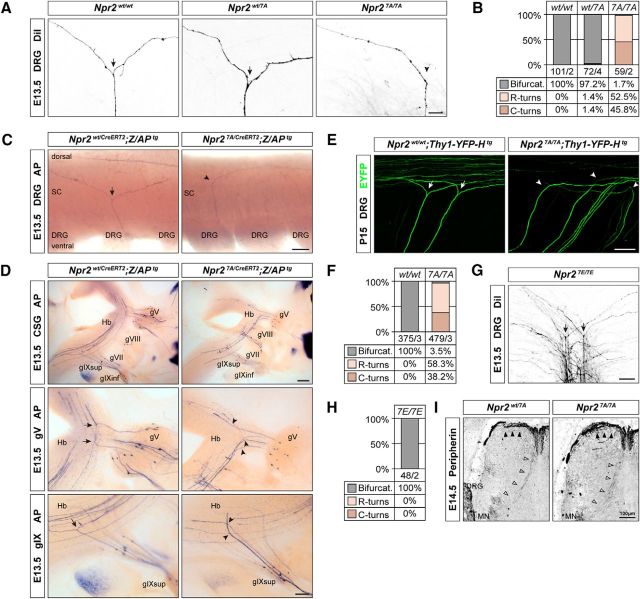
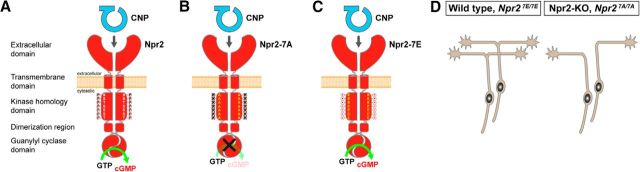
cGMP signaling elicited by activation of the transmembrane receptor guanylyl cyclase Npr2 (also known as guanylyl cyclase B) by the ligand CNP controls sensory axon bifurcation of DRG and cranial sensory ganglion (CSG) neurons entering the spinal cord or hindbrain, respectively. Previous studies have shown that Npr2 is phosphorylated on serine and threonine residues in its kinase homology domain (KHD). However, it is unknown whether phosphorylation of Npr2 is essential for axon bifurcation. Here, we generated a knock-in mouse line in which the seven regulatory serine and threonine residues in the KHD of Npr2 were substituted by alanine (Npr2-7A), resulting in a nonphosphorylatable enzyme. Real-time imaging of cGMP in DRG neurons with a genetically encoded fluorescent cGMP sensor or biochemical analysis of guanylyl cyclase activity in brain or lung tissue revealed the absence of CNP-induced cGMP generation in the Npr27A/7A mutant. Consequently, bifurcation of axons, but not collateral formation, from DRG or CSG in this mouse mutant was perturbed at embryonic and mature stages. In contrast, axon branching was normal in a mouse mutant in which constitutive phosphorylation of Npr2 is mimicked by a replacement of all of the seven serine and threonine sites by glutamic acid (Npr2-7E). Furthermore, we demonstrate that the Npr27A/7A mutation causes dwarfism as described for global Npr2 mutants. In conclusion, our in vivo studies provide strong evidence that phosphorylation of the seven serine and threonine residues in the KHD of Npr2 is an important regulatory element of Npr2-mediated cGMP signaling which affects physiological processes, such as axon bifurcation and bone growth.SIGNIFICANCE STATEMENT The branching of axons is a morphological hallmark of virtually all neurons. It allows an individual neuron to innervate different targets and to communicate with neurons located in different regions of the nervous system. The natriuretic peptide receptor 2 (Npr2), a transmembrane guanylyl cyclase, is essential for the initiation of bifurcation of sensory axons when entering the spinal cord or the hindbrain. By using two genetically engineered mouse lines, we show that phosphorylation of specific serine and threonine residues in juxtamembrane regions of Npr2 are required for its enzymatic activity and for axon bifurcation. These investigations might help to understand the regulation of Npr2 and its integration in intracellular signaling systems.
Keywords: Npr2; axonal branching; cGMP signaling; phosphorylation; sensory axons.
Copyright © 2018 the authors 0270-6474/18/389769-13$15.00/0.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases