

Malic Enzyme 1 (ME1) is pro-oncogenic in ApcMin/+ mice
- PMID: 30250042
- PMCID: PMC6155149
- DOI: 10.1038/s41598-018-32532-w
Malic Enzyme 1 (ME1) is pro-oncogenic in ApcMin/+ mice
Abstract
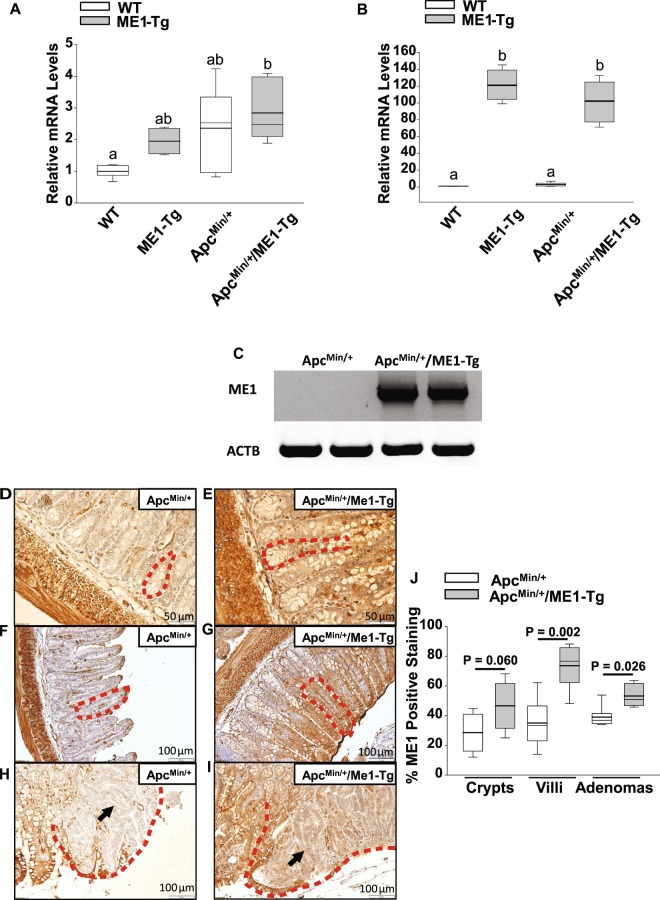
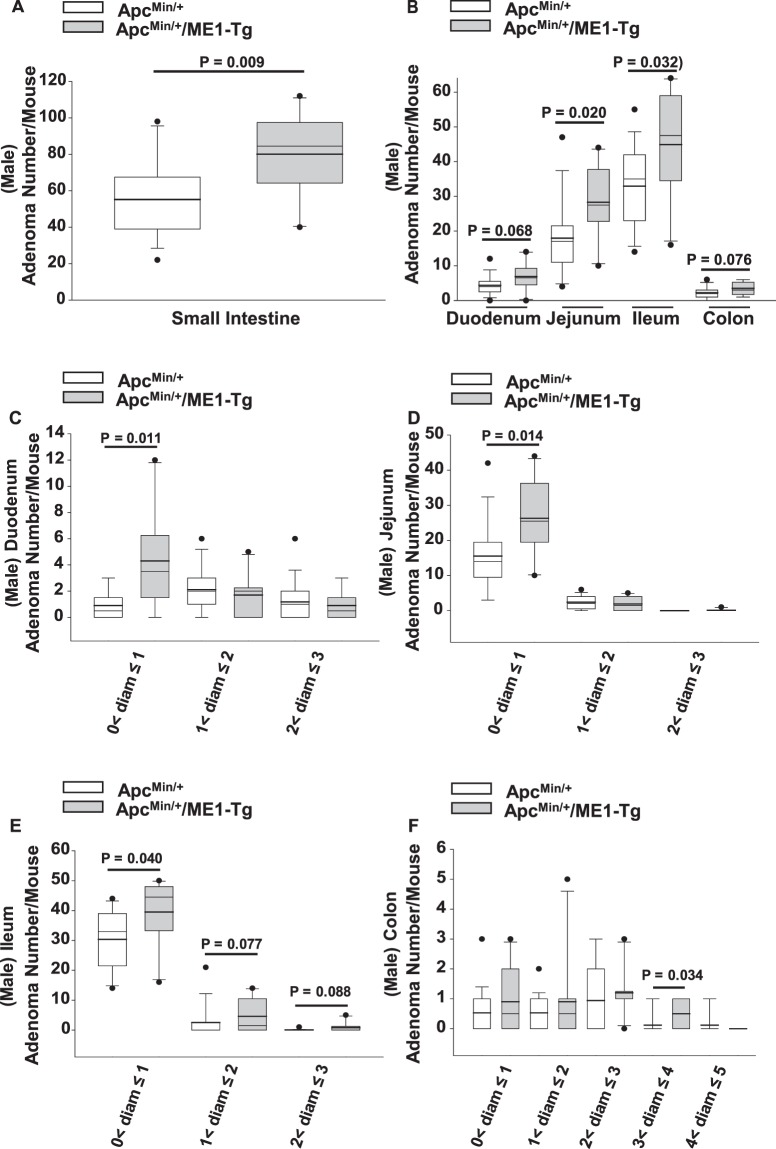
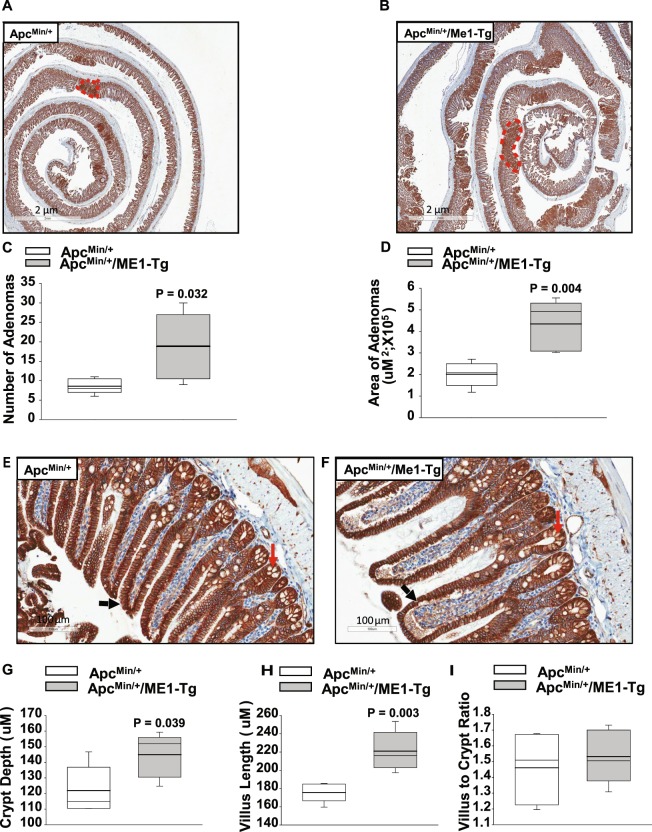
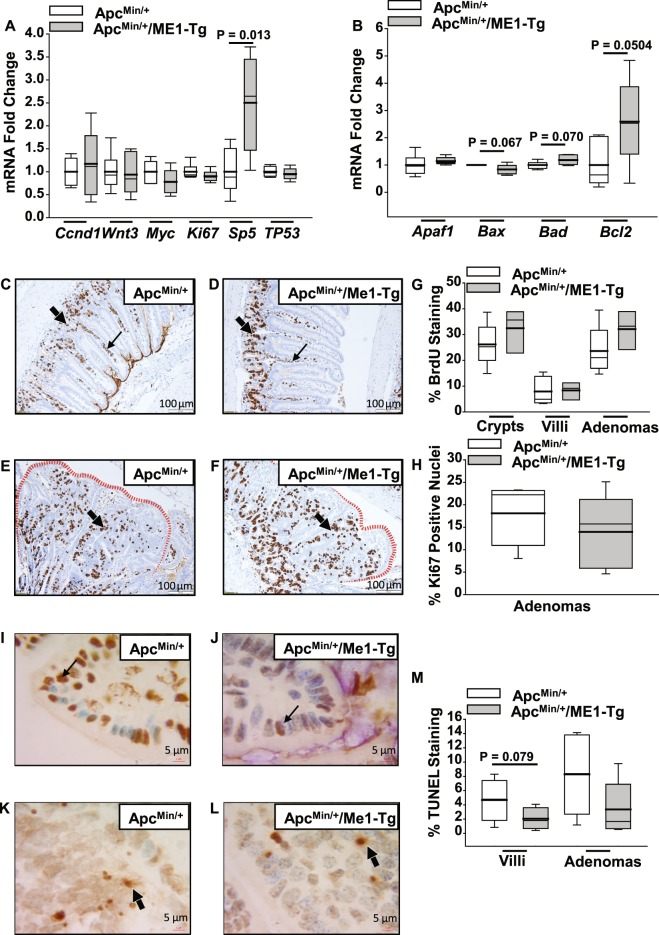
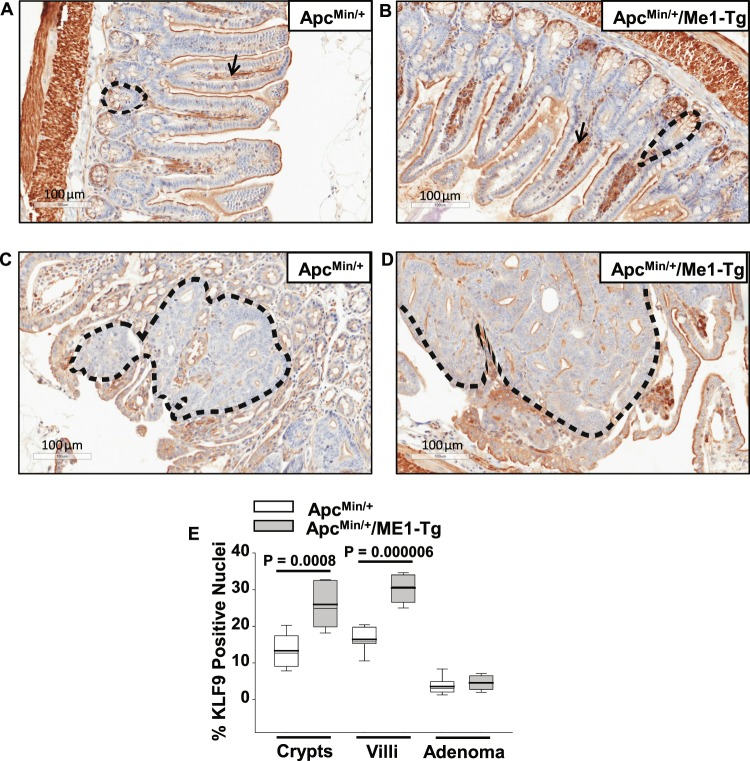
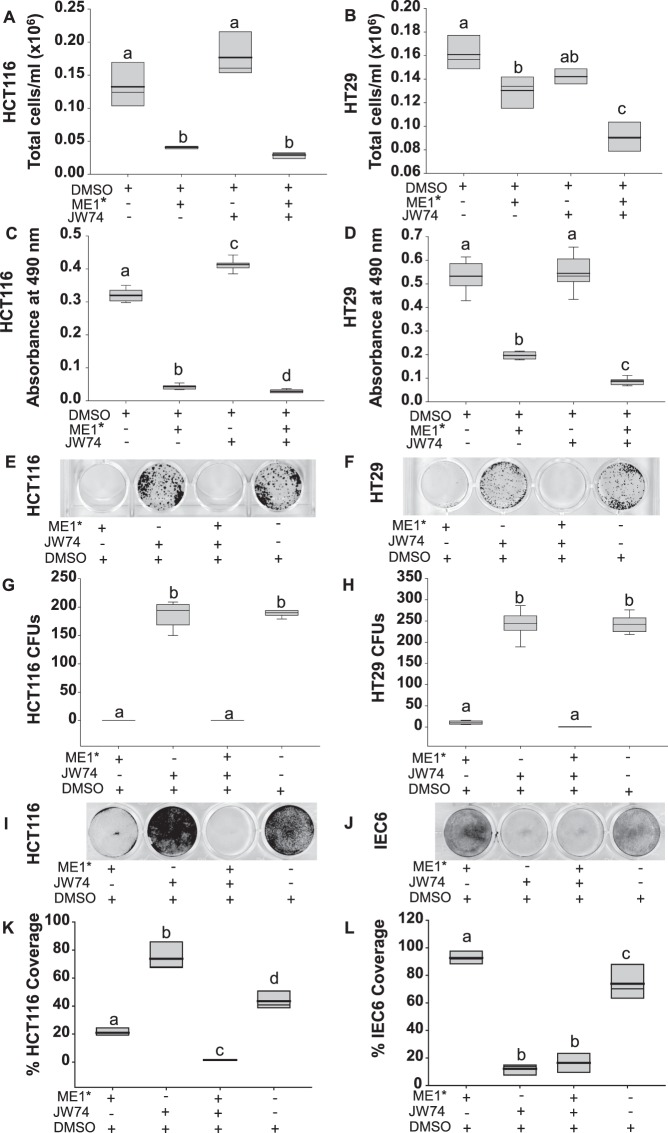
Cytosolic Malic Enzyme (ME1) provides reduced NADP for anabolism and maintenance of redox status. To examine the role of ME1 in tumor genesis of the gastrointestinal tract, we crossed mice having augmented intestinal epithelial expression of ME1 (ME1-Tg mice) with ApcMin/+ mice to obtain male ApcMin/+/ME1-Tg mice. ME1 protein levels were significantly greater within gut epithelium and adenomas of male ApcMin/+/ME1-Tg than ApcMin/+ mice. Male ApcMin/+/ME1-Tg mice had larger and greater numbers of adenomas in the small intestine (jejunum and ileum) than male ApcMin/+ mice. Male ApcMin/+/ME1-Tg mice exhibited greater small intestine crypt depth and villus length in non-adenoma regions, correspondent with increased KLF9 protein abundance in crypts and lamina propria. Small intestines of male ApcMin/+/ME1-Tg mice also had enhanced levels of Sp5 mRNA, suggesting Wnt/β-catenin pathway activation. A small molecule inhibitor of ME1 suppressed growth of human CRC cells in vitro, but had little effect on normal rat intestinal epithelial cells. Targeting of ME1 may add to the armentarium of therapies for cancers of the gastrointestinal tract.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
