

Direct induction of microtubule branching by microtubule nucleation factor SSNA1
- PMID: 30250060
- PMCID: PMC6330057
- DOI: 10.1038/s41556-018-0199-8
Direct induction of microtubule branching by microtubule nucleation factor SSNA1
Abstract
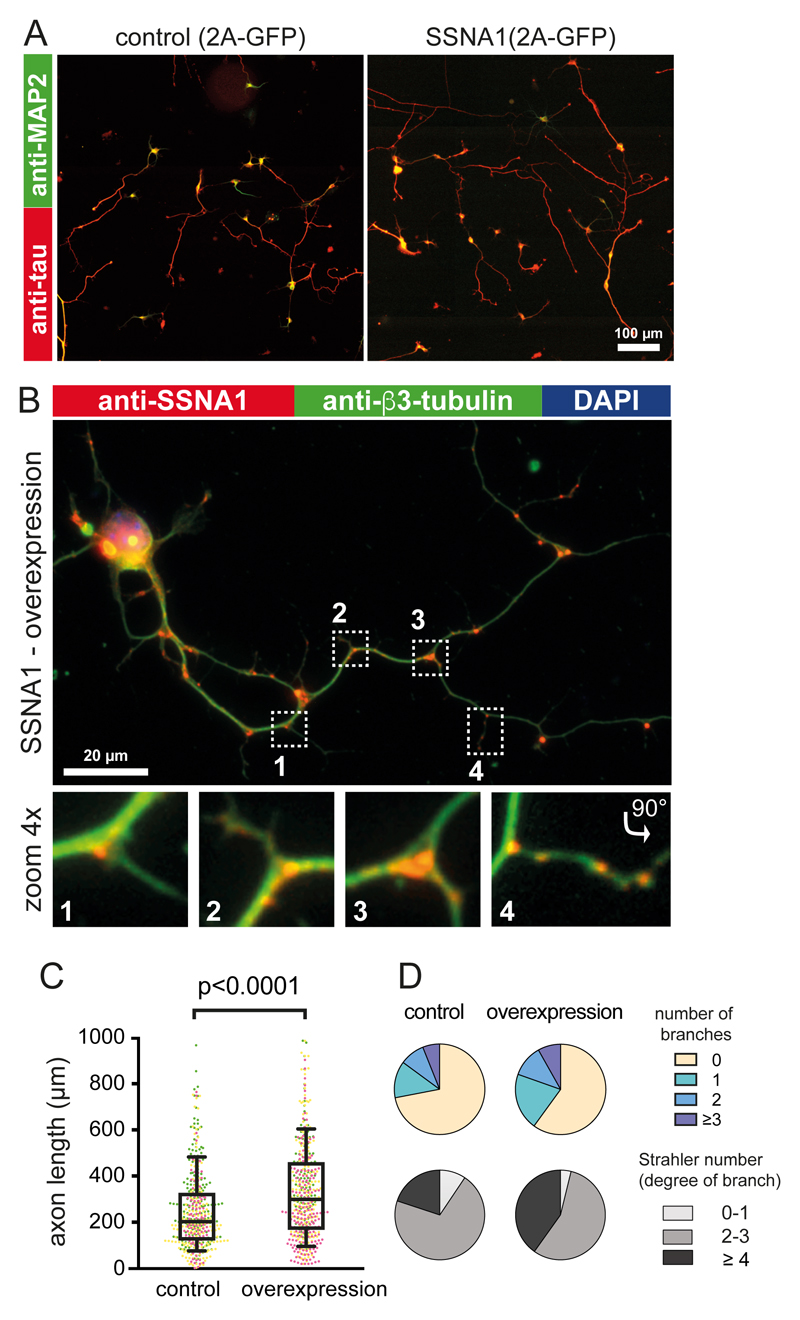
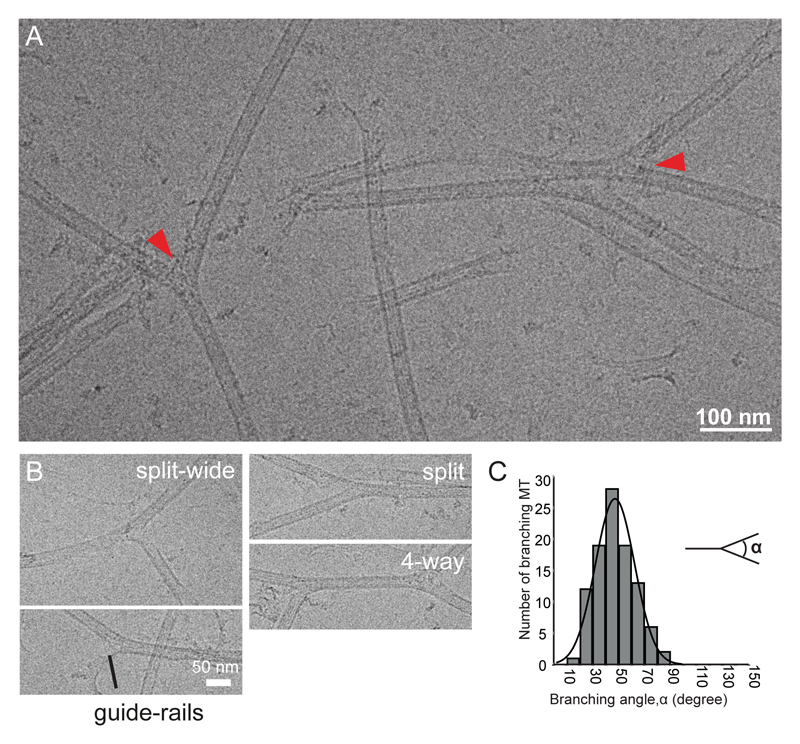
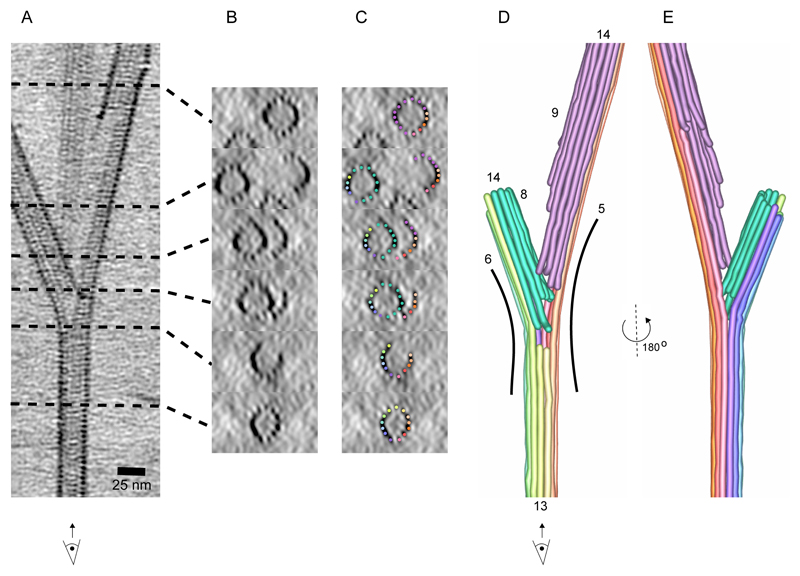
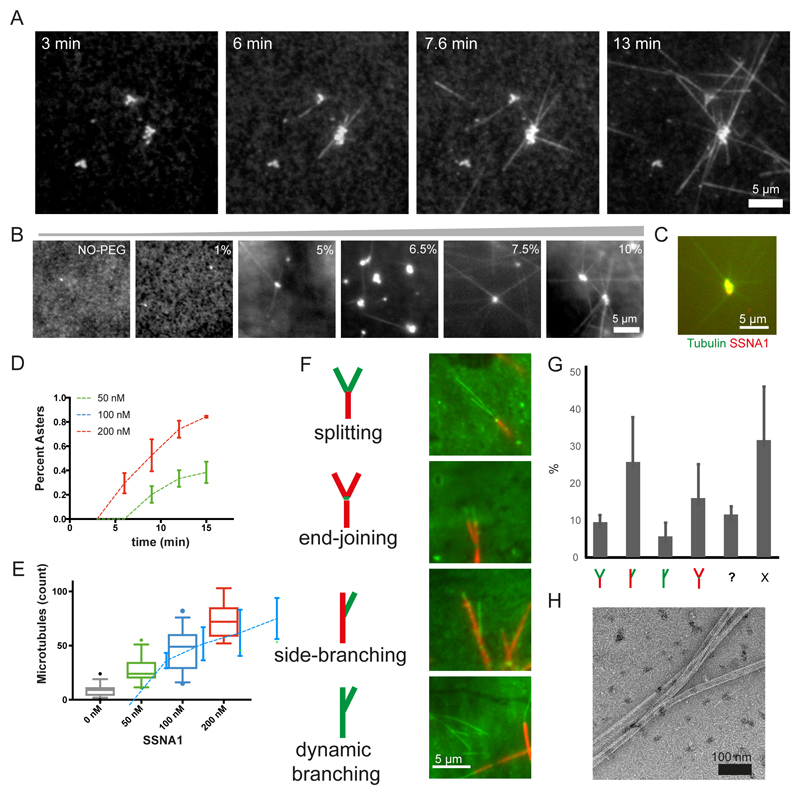
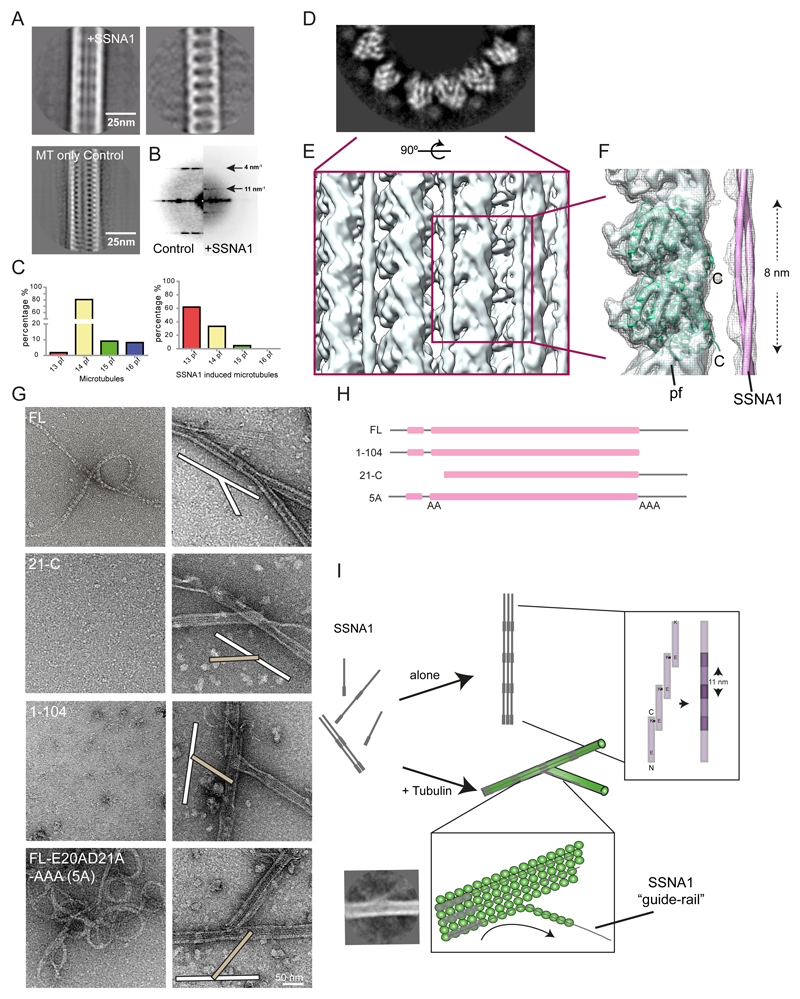
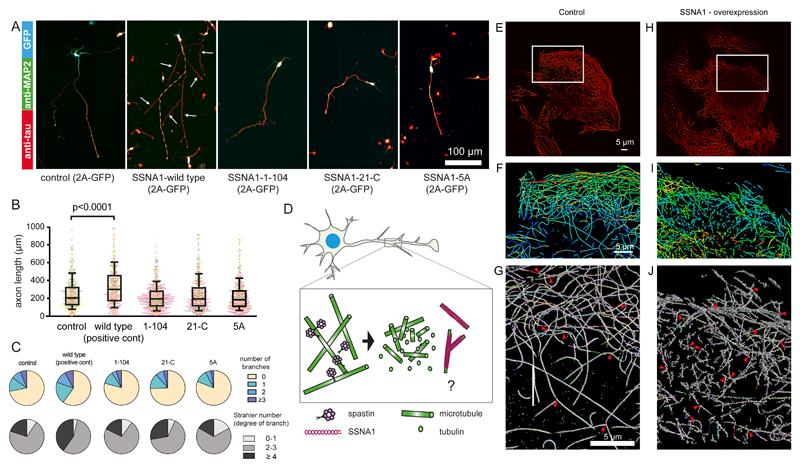
Microtubules are central elements of the eukaryotic cytoskeleton that often function as part of branched networks. Current models for branching include nucleation of new microtubules from severed microtubule seeds or from γ-tubulin recruited to the side of a pre-existing microtubule. Here, we found that microtubules can be directly remodelled into branched structures by the microtubule-remodelling factor SSNA1 (also known as NA14 or DIP13). The branching activity of SSNA1 relies on its ability to self-assemble into fibrils in a head-to-tail fashion. SSNA1 fibrils guide protofilaments of a microtubule to split apart to form daughter microtubules. We further found that SSNA1 localizes at axon branching sites and has a key role in neuronal development. SSNA1 mutants that abolish microtubule branching in vitro also fail to promote axon development and branching when overexpressed in neurons. We have, therefore, discovered a mechanism for microtubule branching and implicated its role in neuronal development.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
