

PAXX and its paralogs synergistically direct DNA polymerase λ activity in DNA repair
- PMID: 30250067
- PMCID: PMC6155126
- DOI: 10.1038/s41467-018-06127-y
PAXX and its paralogs synergistically direct DNA polymerase λ activity in DNA repair
Abstract
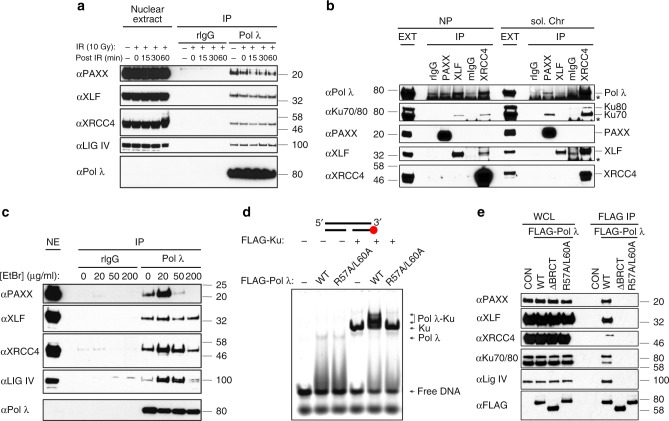
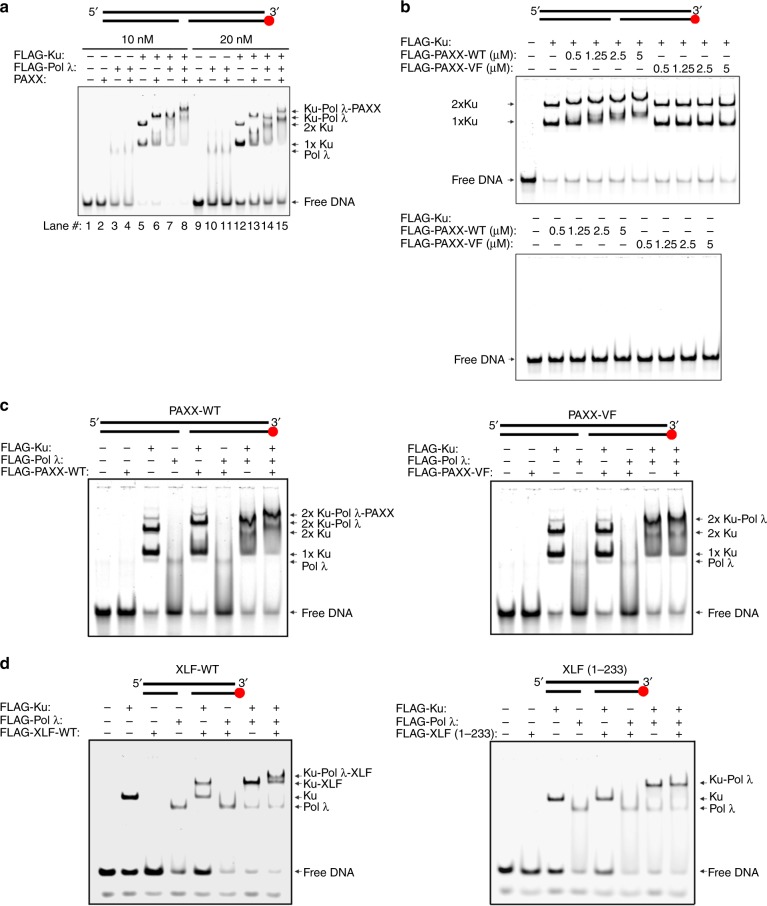
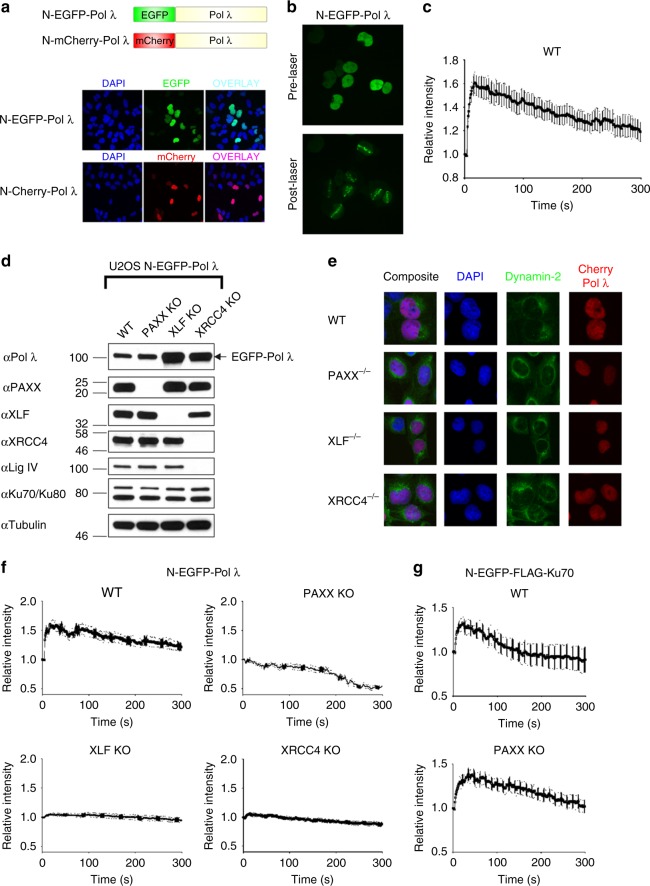
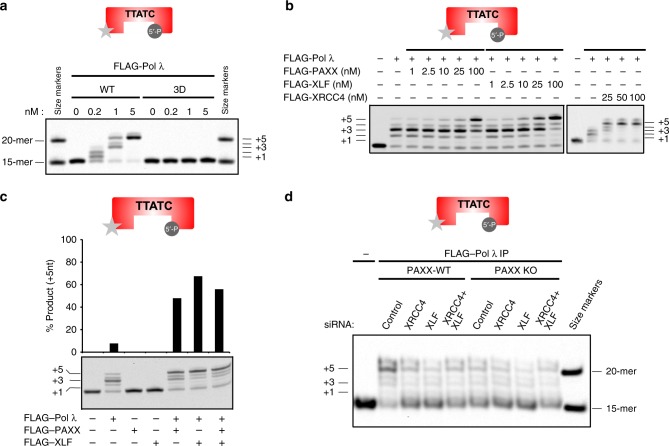
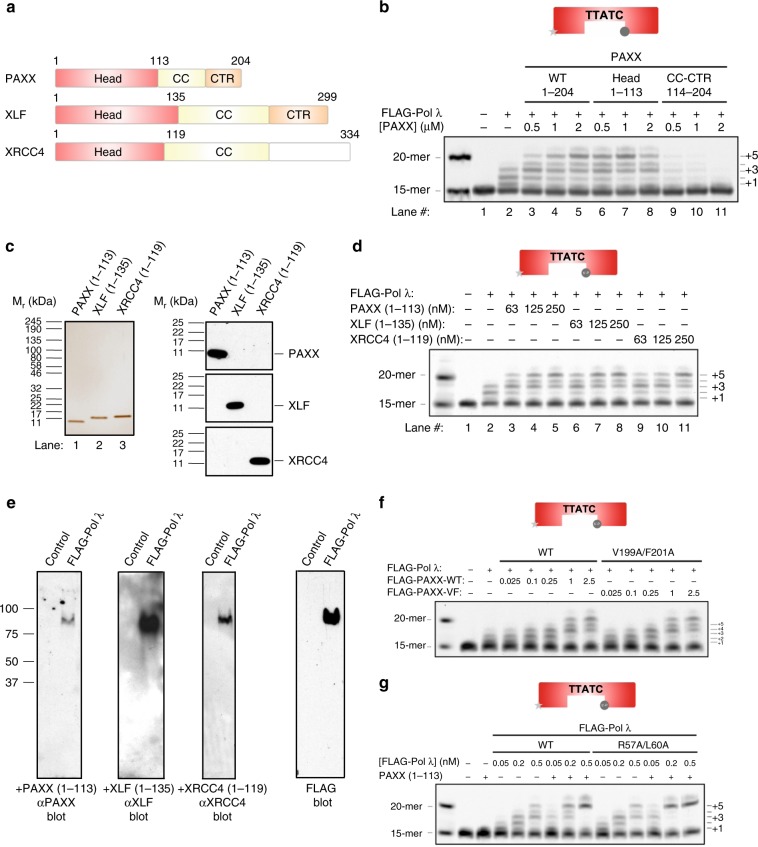
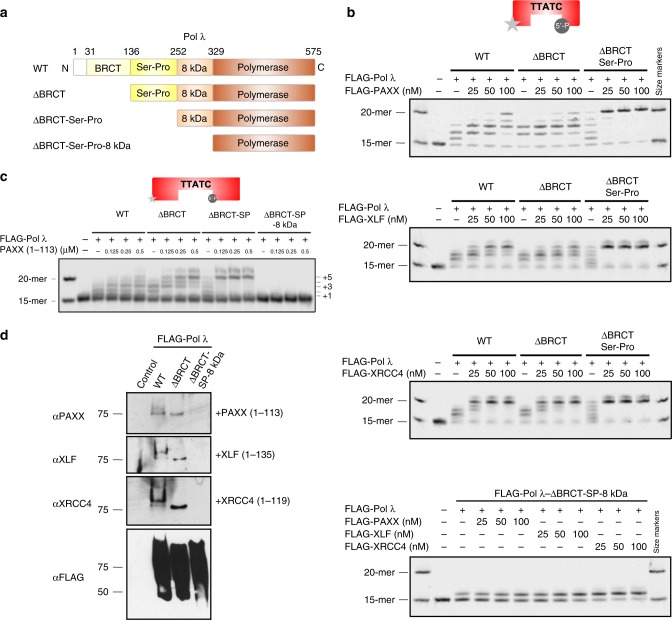
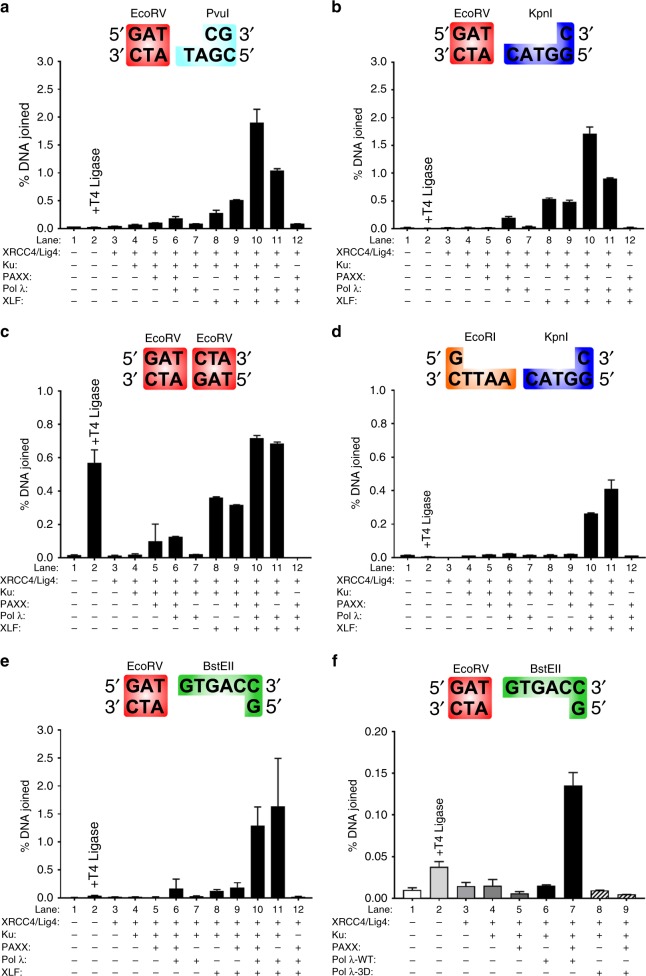
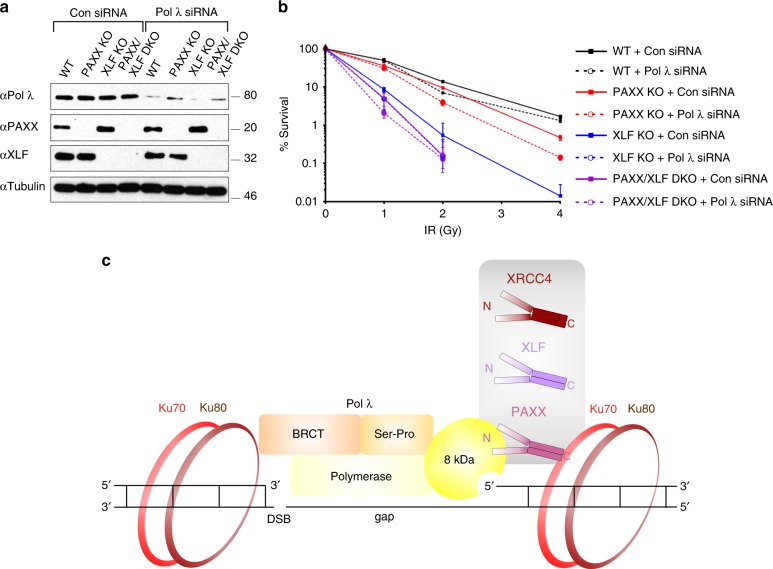
PAXX is a recently identified component of the nonhomologous end joining (NHEJ) DNA repair pathway. The molecular mechanisms of PAXX action remain largely unclear. Here we characterise the interactomes of PAXX and its paralogs, XLF and XRCC4, to show that these factors share the ability to interact with DNA polymerase λ (Pol λ), stimulate its activity and are required for recruitment of Pol λ to laser-induced DNA damage sites. Stimulation of Pol λ activity by XRCC4 paralogs requires a direct interaction between the SP/8 kDa domain of Pol λ and their N-terminal head domains to facilitate recognition of the 5' end of substrate gaps. Furthermore, PAXX and XLF collaborate with Pol λ to promote joining of incompatible DNA ends and are redundant in supporting Pol λ function in vivo. Our findings identify Pol λ as a novel downstream effector of PAXX function and show XRCC4 paralogs act in synergy to regulate polymerase activity in NHEJ.
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Genetic interaction between DNA repair factors PAXX, XLF, XRCC4 and DNA-PKcs in human cells.FEBS Open Bio. 2019 Jul;9(7):1315-1326. doi: 10.1002/2211-5463.12681. Epub 2019 Jun 12. FEBS Open Bio. 2019. PMID: 31141305 Free PMC article.
-
PAXX and XLF DNA repair factors are functionally redundant in joining DNA breaks in a G1-arrested progenitor B-cell line.Proc Natl Acad Sci U S A. 2016 Sep 20;113(38):10619-24. doi: 10.1073/pnas.1611882113. Epub 2016 Sep 6. Proc Natl Acad Sci U S A. 2016. PMID: 27601633 Free PMC article.
-
Absence of XRCC4 and its paralogs in human cells reveal differences in outcomes for DNA repair and V(D)J recombination.DNA Repair (Amst). 2020 Jan;85:102738. doi: 10.1016/j.dnarep.2019.102738. Epub 2019 Nov 12. DNA Repair (Amst). 2020. PMID: 31731258 Free PMC article.
-
Nonhomologous end joining: new accessory factors fine tune the machinery.Trends Genet. 2021 Jun;37(6):582-599. doi: 10.1016/j.tig.2021.03.001. Epub 2021 Mar 27. Trends Genet. 2021. PMID: 33785198 Review.
-
XRCC4 and XLF form long helical protein filaments suitable for DNA end protection and alignment to facilitate DNA double strand break repair.Biochem Cell Biol. 2013 Feb;91(1):31-41. doi: 10.1139/bcb-2012-0058. Epub 2013 Feb 5. Biochem Cell Biol. 2013. PMID: 23442139 Free PMC article. Review.
Cited by
-
Role of Paralogue of XRCC4 and XLF in DNA Damage Repair and Cancer Development.Front Immunol. 2022 Mar 2;13:852453. doi: 10.3389/fimmu.2022.852453. eCollection 2022. Front Immunol. 2022. PMID: 35309348 Free PMC article. Review.
-
Mass spectrometry-based protein‒protein interaction techniques and their applications in studies of DNA damage repair.J Zhejiang Univ Sci B. 2021 Jan 15;22(1):1-20. doi: 10.1631/jzus.B2000356. J Zhejiang Univ Sci B. 2021. PMID: 33448183 Free PMC article. Review.
-
iMUT-seq: high-resolution DSB-induced mutation profiling reveals prevalent homologous-recombination dependent mutagenesis.Nat Commun. 2023 Dec 18;14(1):8419. doi: 10.1038/s41467-023-44167-1. Nat Commun. 2023. PMID: 38110444 Free PMC article.
-
PAXX binding to the NHEJ machinery explains functional redundancy with XLF.Sci Adv. 2023 Jun 2;9(22):eadg2834. doi: 10.1126/sciadv.adg2834. Epub 2023 May 31. Sci Adv. 2023. PMID: 37256950 Free PMC article.
-
PAXX/Ku interaction is rate limiting for repair of double-strand DNA breaks requiring end processing.J Biol Chem. 2025 Aug;301(8):110481. doi: 10.1016/j.jbc.2025.110481. Epub 2025 Jul 12. J Biol Chem. 2025. PMID: 40659092 Free PMC article.
References
-
- Liang S, et al. Achieving selectivity in space and time with DNA double-strand-break response and repair: molecular stages and scaffolds come with strings attached. Struct. Chem. 2017;28:161–171. doi: 10.1007/s11224-016-0841-7. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
