

Structural and functional identification of two distinct inspiratory neuronal populations at the level of the phrenic nucleus in the rat cervical spinal cord
- PMID: 30251026
- PMCID: PMC6373374
- DOI: 10.1007/s00429-018-1757-3
Structural and functional identification of two distinct inspiratory neuronal populations at the level of the phrenic nucleus in the rat cervical spinal cord
Abstract
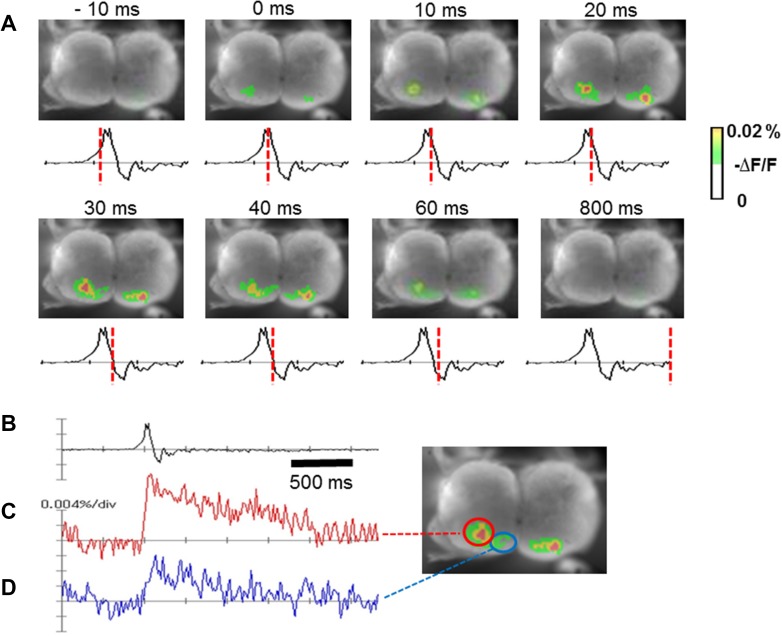
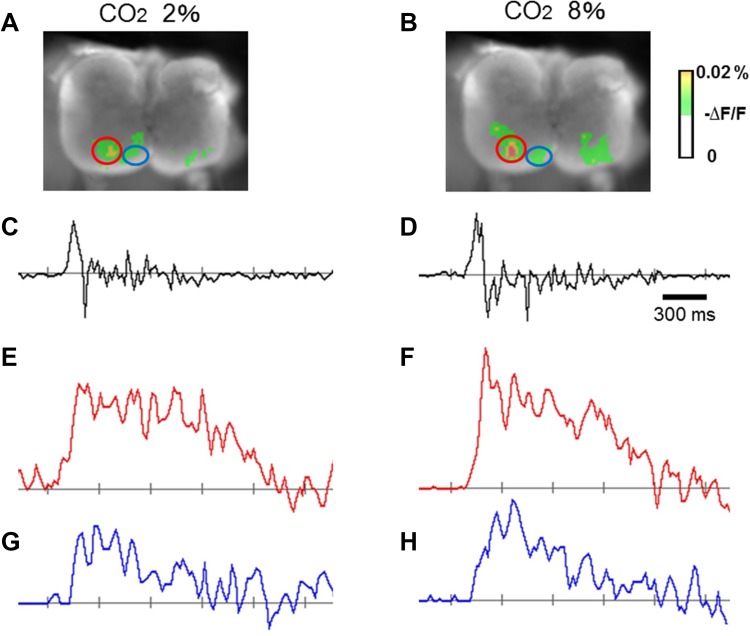
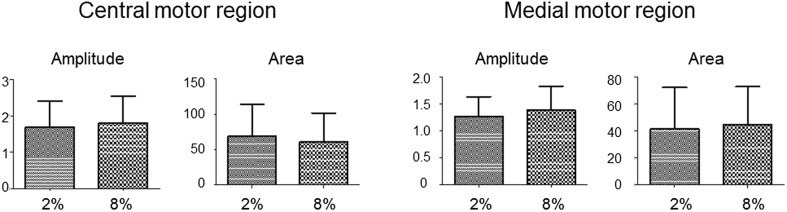
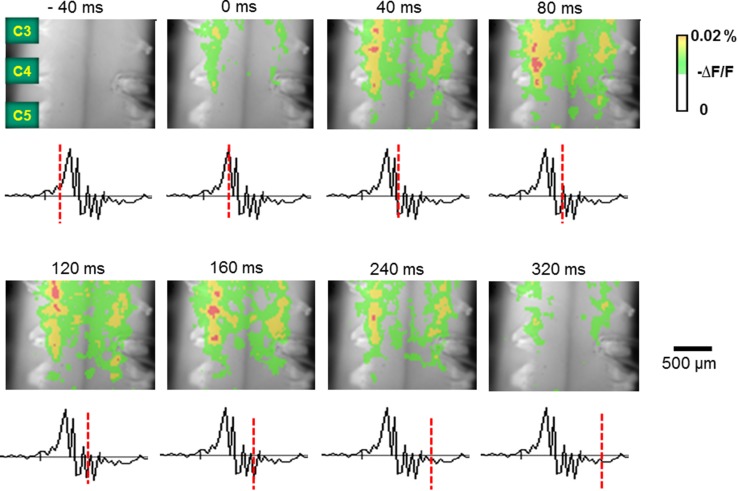
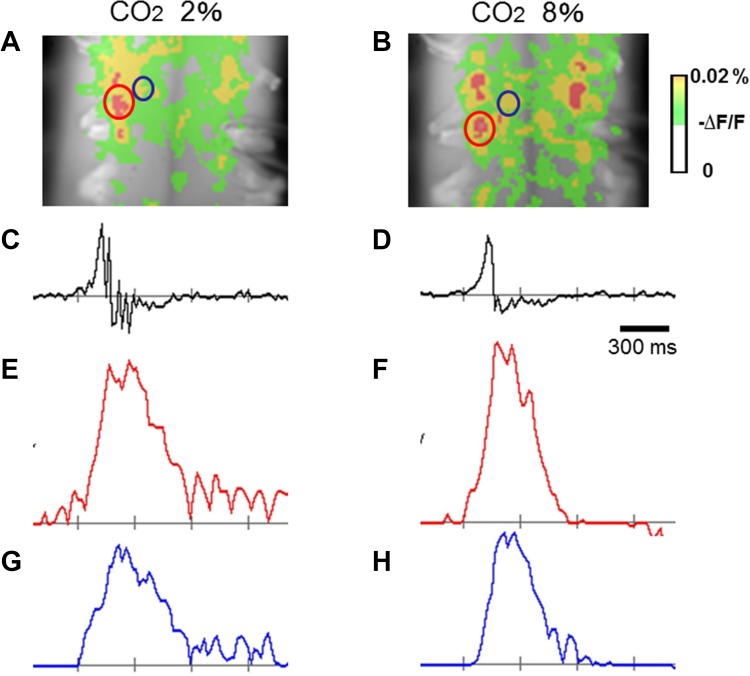
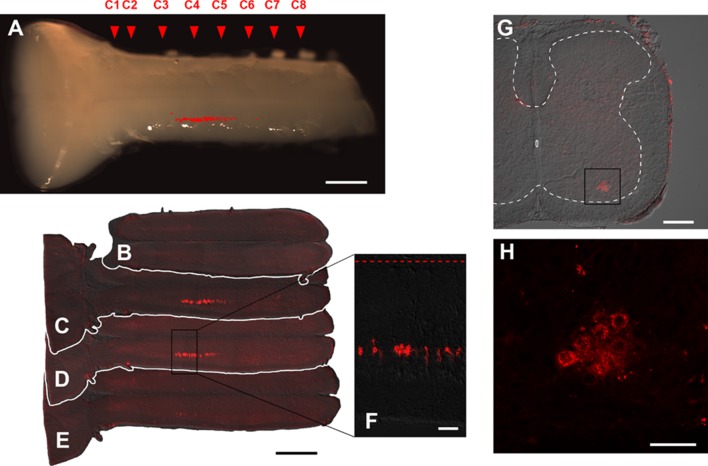
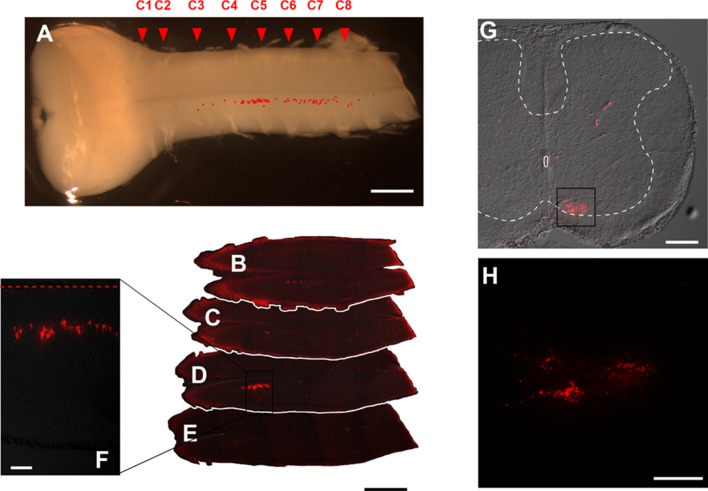
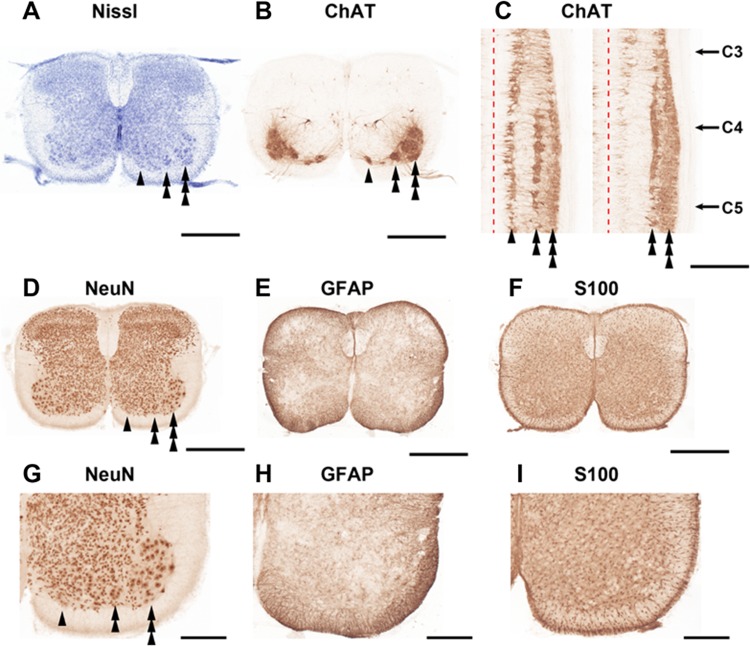
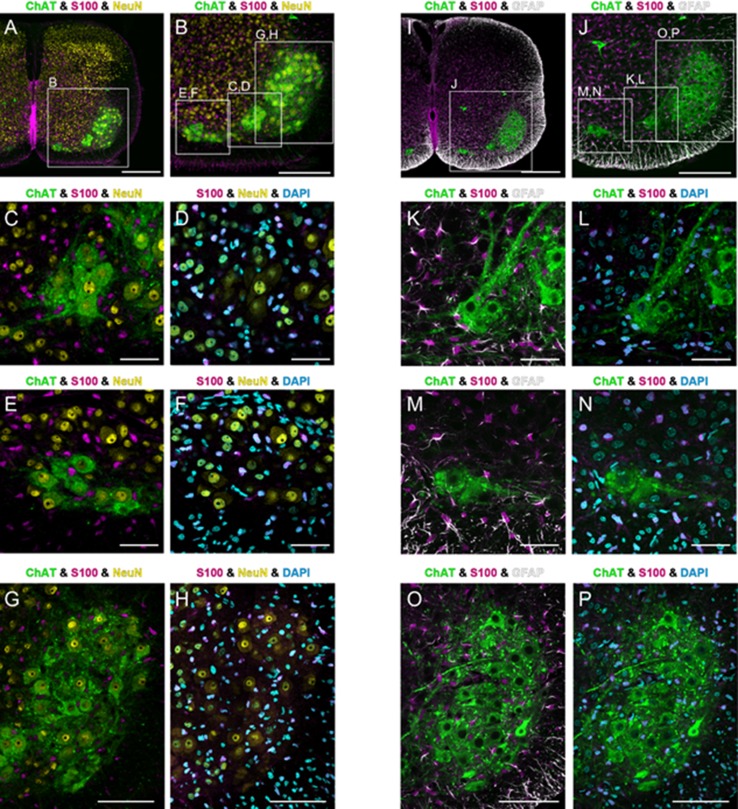
The diaphragm is driven by phrenic motoneurons that are located in the cervical spinal cord. Although the anatomical location of the phrenic nucleus and the function of phrenic motoneurons at a single cellular level have been extensively analyzed, the spatiotemporal dynamics of phrenic motoneuron group activity have not been fully elucidated. In the present study, we analyzed the functional and structural characteristics of respiratory neuron population in the cervical spinal cord at the level of the phrenic nucleus by voltage imaging, together with histological analysis of neuronal and astrocytic distribution in the cervical spinal cord. We found spatially distinct two cellular populations that exhibited synchronized inspiratory activity on the transversely cut plane at C4-C5 levels and on the ventral surface of the mid cervical spinal cord in the isolated brainstem-spinal cord preparation of the neonatal rat. Inspiratory activity of one group emerged in the central portion of the ventral horn that corresponded to the central motor column, and the other appeared in the medial portion of the ventral horn that corresponded to the medial motor column. We identified by retrogradely labeling study that the anatomical distributions of phrenic and scalene motoneurons coincided with optically detected central and medial motor regions, respectively. Furthermore, we anatomically demonstrated closely located features of putative motoneurons, interneurons and astrocytes in these regions. Collectively, we report that phrenic and scalene motoneuron populations show synchronized inspiratory activities with distinct anatomical locations in the mid cervical spinal cord.
Keywords: Astrocyte; Cervical spinal cord; Interneuron; Phrenic motoneuron; Phrenic nucleus; Respiratory control; Scalene motoneuron; Voltage imaging.
Conflict of interest statement
Conflict of interest
The authors declare no competing financial interests.
Ethical approval
All experiments were carried out in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 80-23) revised 1996 and with the Guiding Principles for the Care and Use of Animals of the Physiological Society of Japan. Experiments for voltage imaging and for anatomical analysis were approved by the Animal Experiment Ethics Committees of Keio University (Permit Number: 020062) and Shimane University (Permit Numbers: 03-34, H17-7, H19-53, H20-32 and IZ25-14).
Informed consent
Not applicable.
Figures
Similar articles
-
Phrenic motor outputs in response to bronchopulmonary C-fibre activation following chronic cervical spinal cord injury.J Physiol. 2016 Oct 15;594(20):6009-6024. doi: 10.1113/JP272287. Epub 2016 Jun 3. J Physiol. 2016. PMID: 27106483 Free PMC article.
-
Brainstem network controlling descending drive to phrenic motoneurons in rat.J Comp Neurol. 1994 Sep 1;347(1):64-86. doi: 10.1002/cne.903470106. J Comp Neurol. 1994. PMID: 7798382
-
Coupling multielectrode array recordings with silver labeling of recording sites to study cervical spinal network connectivity.J Neurophysiol. 2017 Mar 1;117(3):1014-1029. doi: 10.1152/jn.00638.2016. Epub 2016 Dec 14. J Neurophysiol. 2017. PMID: 27974450 Free PMC article.
-
Pre-phrenic interneurons: Characterization and role in phrenic pattern formation and respiratory recovery following spinal cord injury.Respir Physiol Neurobiol. 2019 Jul;265:24-31. doi: 10.1016/j.resp.2018.09.005. Epub 2018 Oct 10. Respir Physiol Neurobiol. 2019. PMID: 30315961 Review.
-
Functional recovery after cervical spinal cord injury: Role of neurotrophin and glutamatergic signaling in phrenic motoneurons.Respir Physiol Neurobiol. 2016 Jun;226:128-36. doi: 10.1016/j.resp.2015.10.009. Epub 2015 Oct 23. Respir Physiol Neurobiol. 2016. PMID: 26506253 Free PMC article. Review.
Cited by
-
The integrated brain network that controls respiration.Elife. 2023 Mar 8;12:e83654. doi: 10.7554/eLife.83654. Elife. 2023. PMID: 36884287 Free PMC article. Review.
-
Proposal for Manual Osteopathic Treatment of the Phrenic Nerve.Cureus. 2024 Apr 11;16(4):e58012. doi: 10.7759/cureus.58012. eCollection 2024 Apr. Cureus. 2024. PMID: 38606024 Free PMC article. Review.
-
Sex- and Region-Specific Differences in the Transcriptomes of Rat Microglia from the Brainstem and Cervical Spinal Cord.J Pharmacol Exp Ther. 2020 Oct;375(1):210-222. doi: 10.1124/jpet.120.266171. Epub 2020 Jul 13. J Pharmacol Exp Ther. 2020. PMID: 32661056 Free PMC article.
-
Structural and functional connectivity from the dorsomedial hypothalamus to the ventral medulla as a chronological amplifier of sympathetic outflow.Sci Rep. 2020 Aug 7;10(1):13325. doi: 10.1038/s41598-020-70234-4. Sci Rep. 2020. PMID: 32770006 Free PMC article.
-
Central Autonomic Mechanisms Involved in the Control of Laryngeal Activity and Vocalization.Biology (Basel). 2024 Feb 13;13(2):118. doi: 10.3390/biology13020118. Biology (Basel). 2024. PMID: 38392336 Free PMC article. Review.
References
-
- Allan DW, Greer JJ. Development of phrenic motoneuron morphology in the fetal rat. J Comp Neurol. 1997;382:469–479. - PubMed
-
- Anderson WJ, Bellinger DL, Lorton D. Morphology of dendrite bundles in the cervical spinal cord of the rat: a light microscopic study. Exp Neurol. 1988;100:121–138. - PubMed
-
- Aoyama R, Okada Y, Yokota S, Yasui Y, Fukuda K, Shinozaki Y, Yoshida H, Nakamura M, Chiba K, Yasui Y, Kato F, Toyama Y. Spatiotemporal and anatomical analyses of P2X receptor-mediated neuronal and glial processing of sensory signals in the rat dorsal horn. Pain. 2011;152:2085–2097. - PubMed
-
- Barber RP, Phelps PE, Houser CR, Crawford GD, Salvaterra PM, Vaughn JE. The morphology and distribution of neurons containing choline acetyltransferase in the adult rat spinal cord: an immunocytochemical study. J Comp Neurol. 1984;229(3):329–346. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
