

Insertion sequence 1 from calpain-3 is functional in calpain-2 as an internal propeptide
- PMID: 30254072
- PMCID: PMC6240860
- DOI: 10.1074/jbc.RA118.004803
Insertion sequence 1 from calpain-3 is functional in calpain-2 as an internal propeptide
Abstract
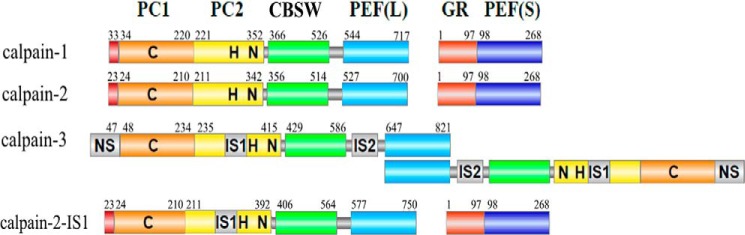
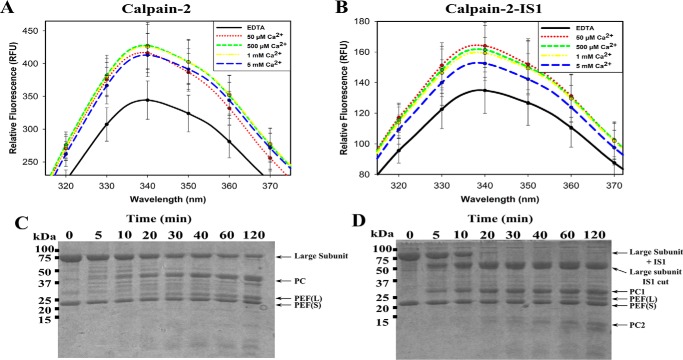
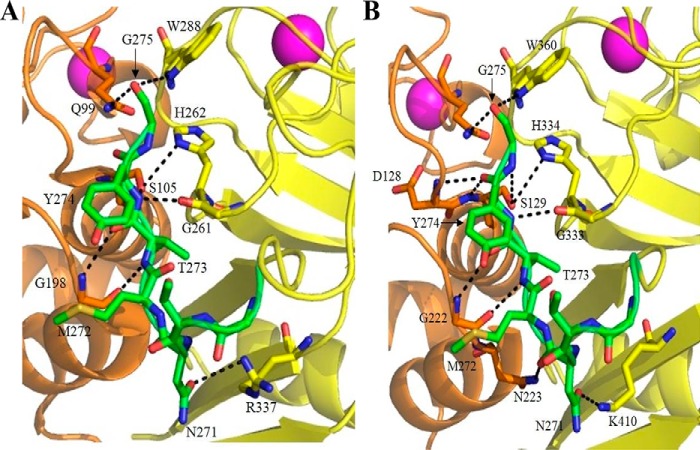
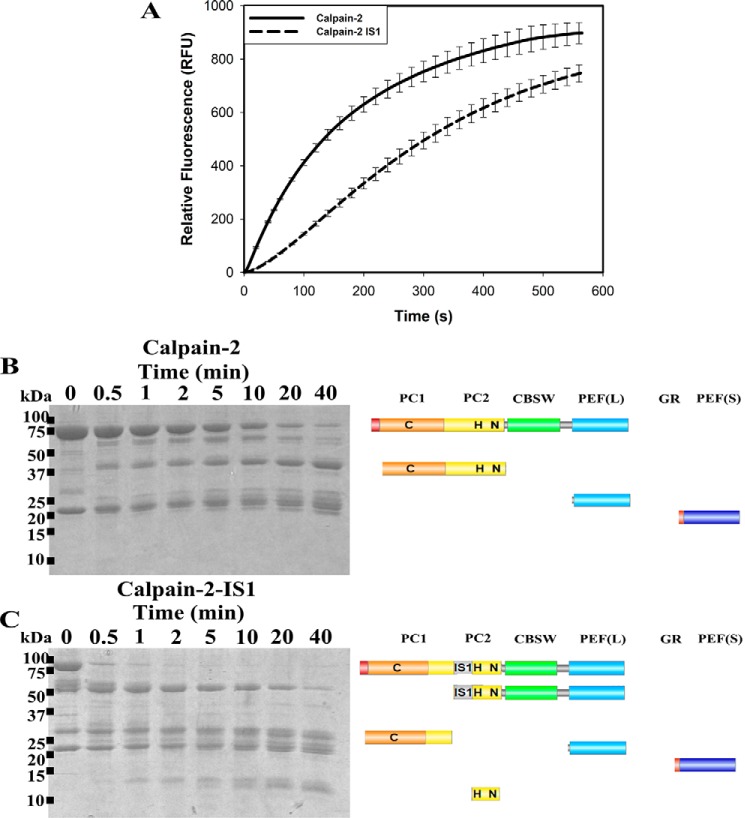
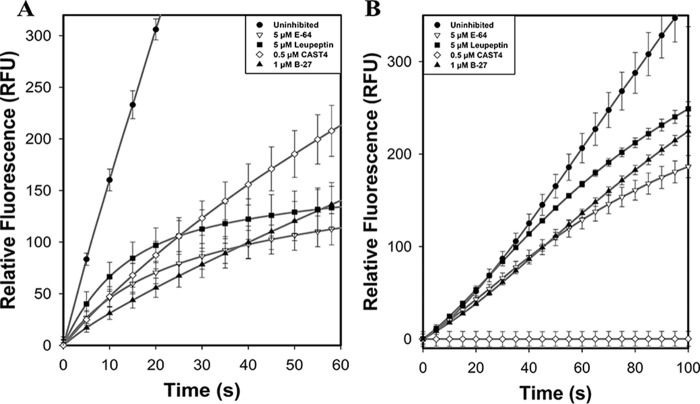
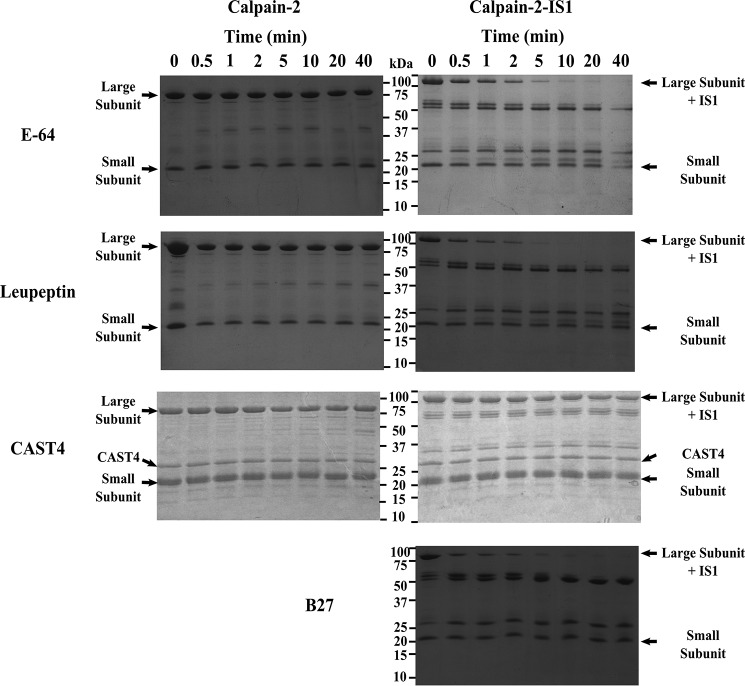
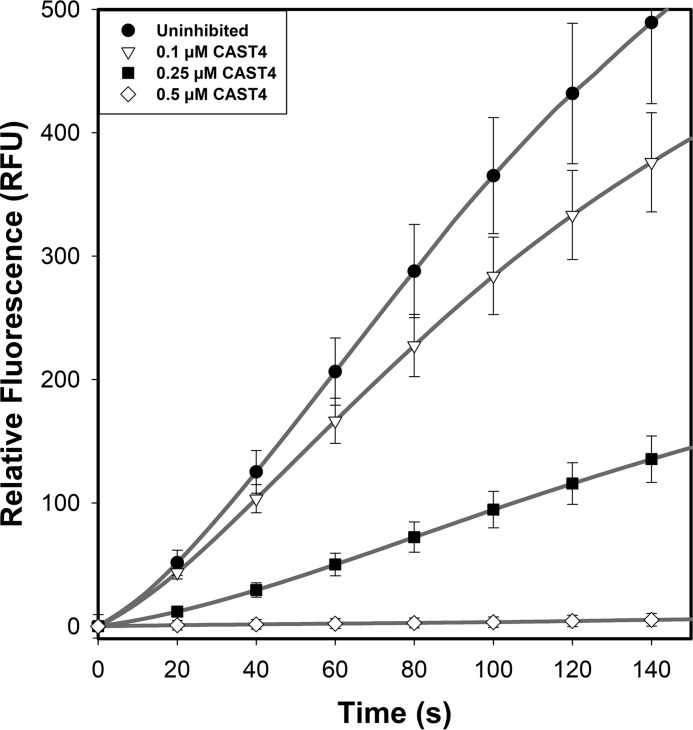
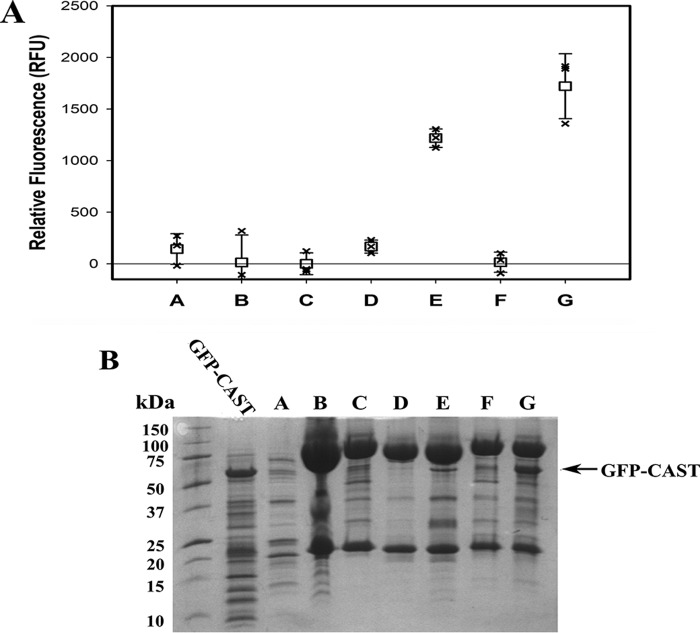
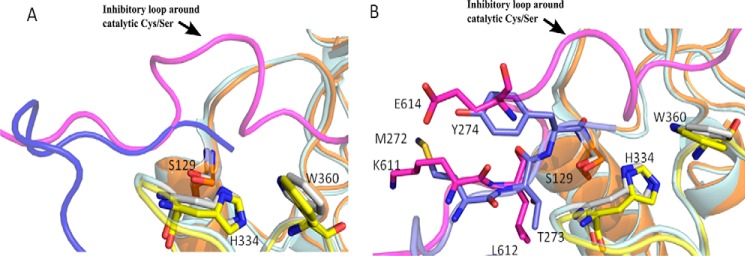
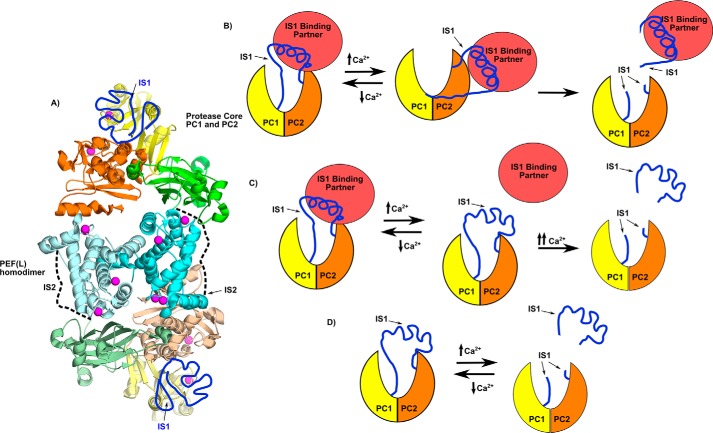
Calpains are intracellular, calcium-activated cysteine proteases. Calpain-3 is abundant in skeletal muscle, where its mutation-induced loss of function causes limb-girdle muscular dystrophy type 2A. Unlike the small subunit-containing calpain-1 and -2, the calpain-3 isoform homodimerizes through pairing of its C-terminal penta-EF-hand domain. It also has two unique insertion sequences (ISs) not found in the other calpains: IS1 within calpain-3's protease core and IS2 just prior to the penta-EF-hand domain. Production of either native or recombinant full-length calpain-3 to characterize the function of these ISs is challenging. Therefore, here we used recombinant rat calpain-2 as a stable surrogate and inserted IS1 into its equivalent position in the protease core. As it does in calpain-3, IS1 occupied the catalytic cleft and restricted the enzyme's access to substrate and inhibitors. Following activation by Ca2+, IS1 was rapidly cleaved by intramolecular autolysis, permitting the enzyme to freely accept substrate and inhibitors. The surrogate remained functional until extensive intermolecular autoproteolysis inactivated the enzyme, as is typical of calpain-2. Although the small-molecule inhibitors E-64 and leupeptin limited intermolecular autolysis of the surrogate, they did not block the initial intramolecular cleavage of IS1, establishing its role as a propeptide. Surprisingly, the large-molecule calpain inhibitor, calpastatin, completely blocked enzyme activity, even with IS1 intact. We suggest that calpastatin is large enough to oust IS1 from the catalytic cleft and take its place. We propose an explanation for why calpastatin can inhibit calpain-2 bearing the IS1 insertion but cannot inhibit WT calpain-3.
Keywords: calpain; calpain inhibitor; calpain-2; calpain-3/p94; calpastatin; cysteine protease; insertion sequence 1 (IS1); limb-girdle muscular dystrophy; muscular dystrophy; p94; propeptide; protease; protease inhibitor; protein engineering; structure-function.
© 2018 McCartney et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
