

Corepressor SMRT is required to maintain Hox transcriptional memory during somitogenesis
- PMID: 30254164
- PMCID: PMC6187131
- DOI: 10.1073/pnas.1809480115
Corepressor SMRT is required to maintain Hox transcriptional memory during somitogenesis
Abstract
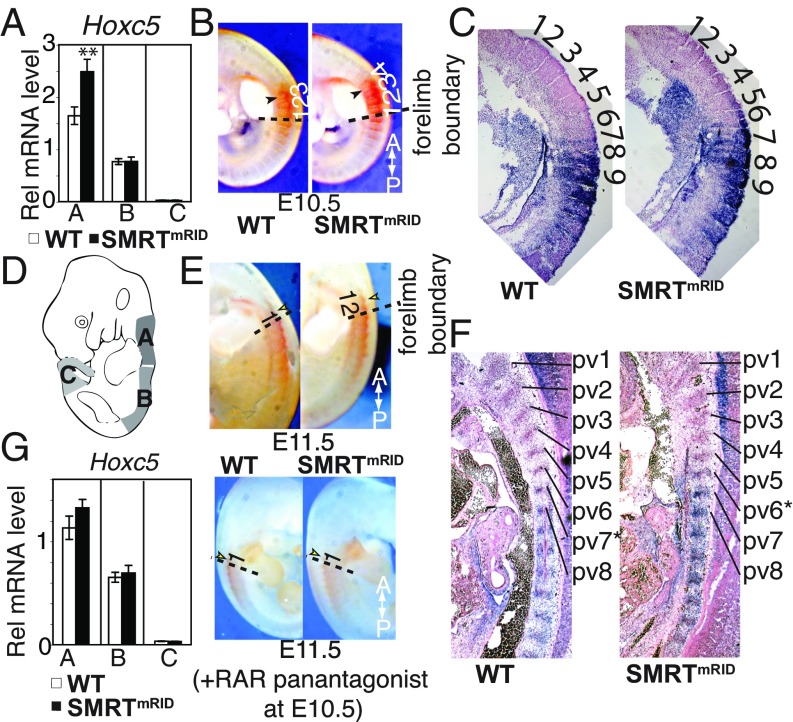
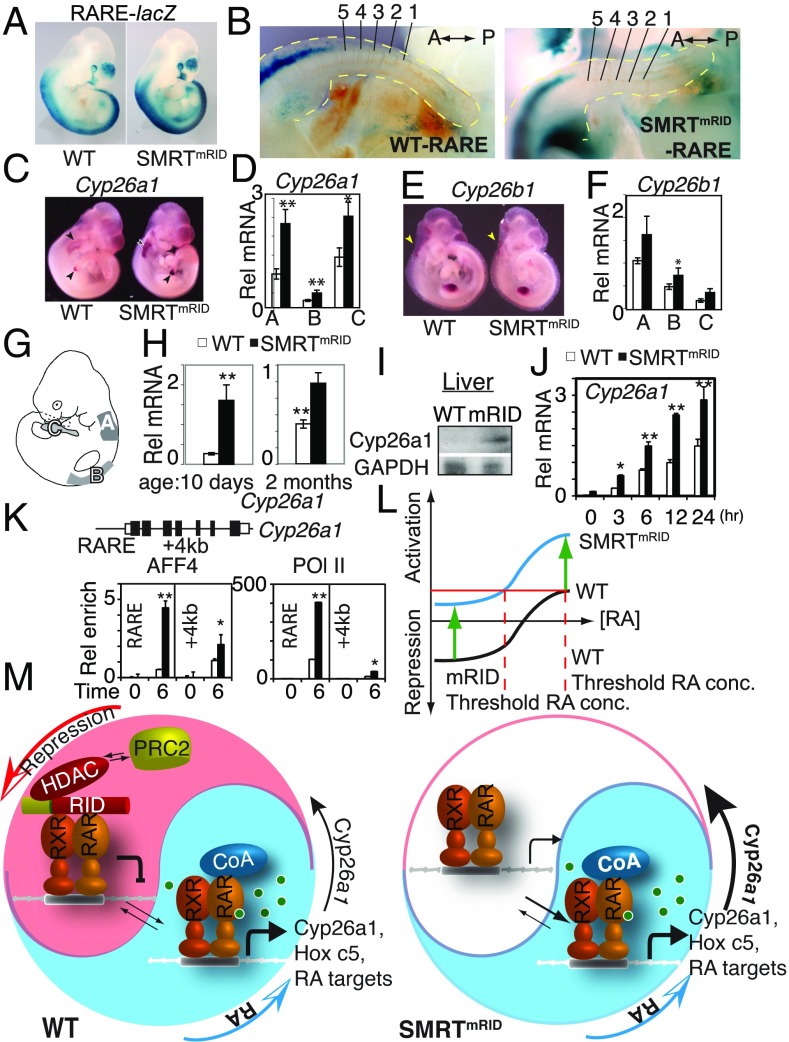
Nuclear hormone receptors (NRs), such as retinoic acid receptors (RARs), play critical roles in vertebrate development and homeostasis by regulating target gene transcription. Their activity is controlled by ligand-dependent release of corepressors and subsequent recruitment of coactivators, but how these individual receptor modes contribute to development are unknown. Here, we show that mice carrying targeted knockin mutations in the corepressor Silencing Mediator of Retinoid and Thyroid hormone receptor (SMRT) that specifically disable SMRT function in NR signaling (SMRTmRID), display defects in cranial neural crest cell-derived structures and posterior homeotic transformations of axial vertebrae. SMRTmRID embryos show enhanced transcription of RAR targets including Hox loci, resulting in respecification of vertebral identities. Up-regulated histone acetylation and decreased H3K27 methylation are evident in the Hox loci whose somitic expression boundaries are rostrally shifted. Furthermore, enhanced recruitment of super elongation complex is evident in rapidly induced non-Pol II-paused targets in SMRTmRID embryonic stem cells. These results demonstrate that SMRT-dependent repression of RAR is critical to establish and maintain the somitic Hox code and segmental identity during fetal development via epigenetic marking of target loci.
Keywords: SMRT; homeotic transformation; retinoic acid receptor; somitogenesis; transcriptional repression.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Chen JD, Evans RM. A transcriptional co-repressor that interacts with nuclear hormone receptors. Nature. 1995;377:454–457. - PubMed
-
- Hörlein AJ, et al. Ligand-independent repression by the thyroid hormone receptor mediated by a nuclear receptor co-repressor. Nature. 1995;377:397–404. - PubMed
-
- Lonard DM, O’malley BW. Nuclear receptor coregulators: Judges, juries, and executioners of cellular regulation. Mol Cell. 2007;27:691–700. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
