

Structure of a PSI-LHCI-cyt b6f supercomplex in Chlamydomonas reinhardtii promoting cyclic electron flow under anaerobic conditions
- PMID: 30254175
- PMCID: PMC6187136
- DOI: 10.1073/pnas.1809973115
Structure of a PSI-LHCI-cyt b6f supercomplex in Chlamydomonas reinhardtii promoting cyclic electron flow under anaerobic conditions
Abstract
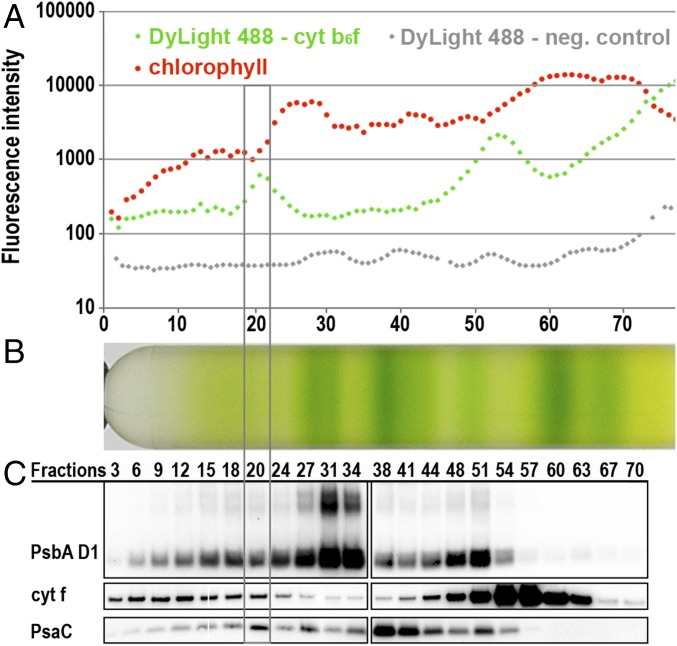
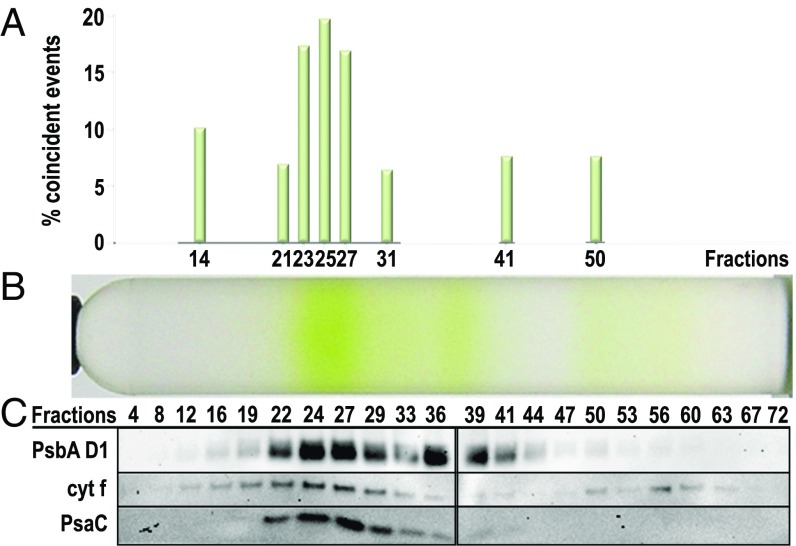
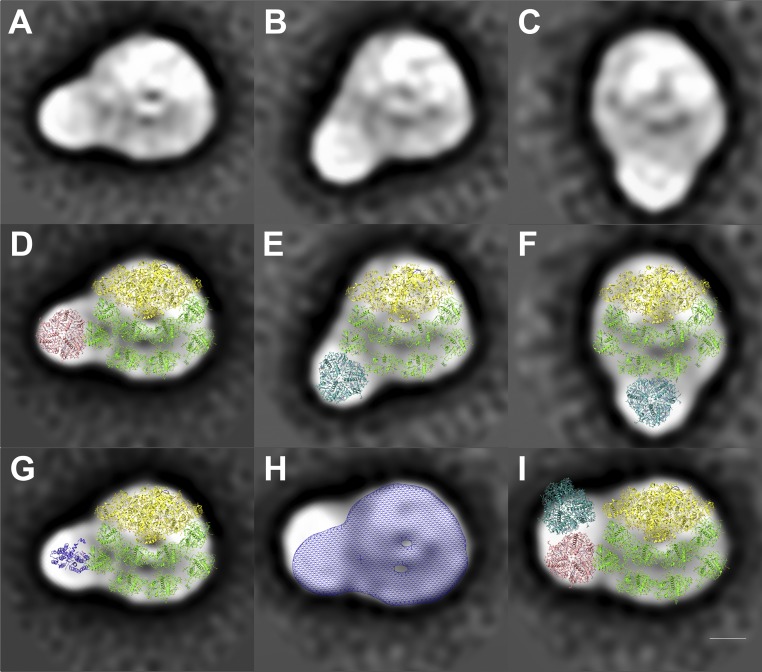
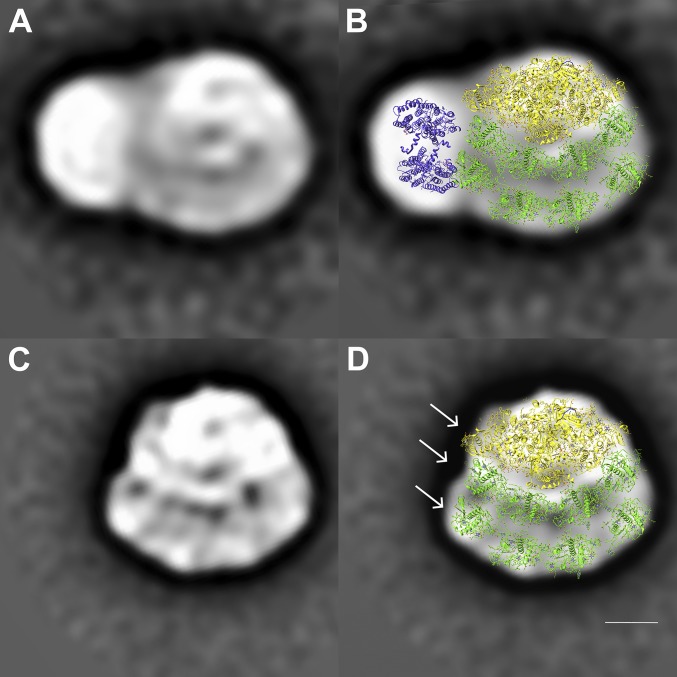
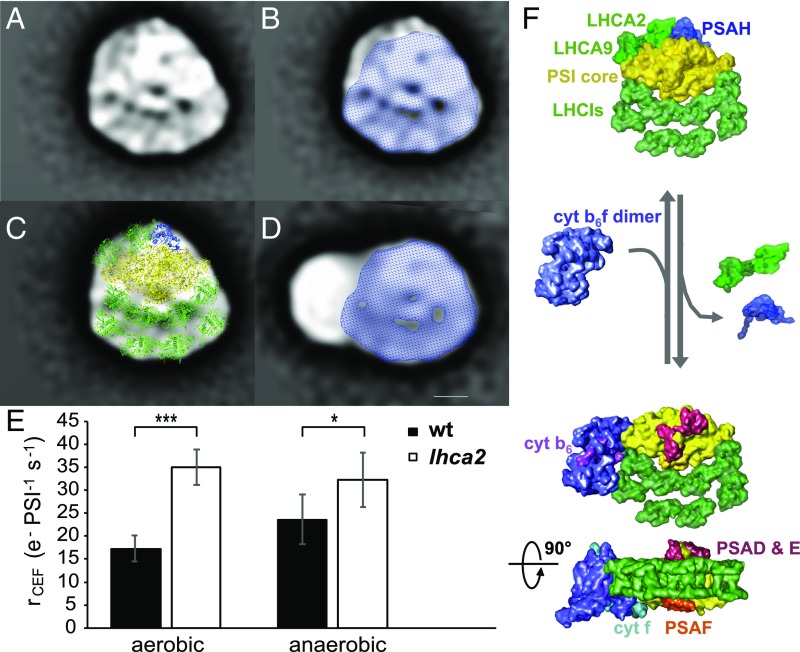
Photosynthetic linear electron flow (LEF) produces ATP and NADPH, while cyclic electron flow (CEF) exclusively drives photophosphorylation to supply extra ATP. The fine-tuning of linear and cyclic electron transport levels allows photosynthetic organisms to balance light energy absorption with cellular energy requirements under constantly changing light conditions. As LEF and CEF share many electron transfer components, a key question is how the same individual structural units contribute to these two different functional modes. Here, we report the structural identification of a photosystem I (PSI)-light harvesting complex I (LHCI)-cytochrome (cyt) b6f supercomplex isolated from the unicellular alga Chlamydomonas reinhardtii under anaerobic conditions, which induces CEF. This provides strong evidence for the model that enhanced CEF is induced by the formation of CEF supercomplexes, when stromal electron carriers are reduced, to generate additional ATP. The additional identification of PSI-LHCI-LHCII complexes is consistent with recent findings that both CEF enhancement and state transitions are triggered by similar conditions, but can occur independently from each other. Single molecule fluorescence correlation spectroscopy indicates a physical association between cyt b6f and fluorescent chlorophyll containing PSI-LHCI supercomplexes. Single particle analysis identified top-view projections of the corresponding PSI-LHCI-cyt b6f supercomplex. Based on molecular modeling and mass spectrometry analyses, we propose a model in which dissociation of LHCA2 and LHCA9 from PSI supports the formation of this CEF supercomplex. This is supported by the finding that a Δlhca2 knockout mutant has constitutively enhanced CEF.
Keywords: Chlamydomonas reinhardtii; cyclic electron flow; cytochrome b6f; photosystem I; supercomplex.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Arnon DI, Allen MB, Whatley FR. Photosynthesis by isolated chloroplasts. Nature. 1954;174:394–396. - PubMed
-
- Bassham JA, Benson AA, Calvin M. The path of carbon in photosynthesis. J Biol Chem. 1950;185:781–787. - PubMed
-
- Calvin M, Benson AA. The path of carbon in photosynthesis. Science. 1948;107:476–480. - PubMed
-
- Allen JF. Cyclic, pseudocyclic and noncyclic photophosphorylation: New links in the chain. Trends Plant Sci. 2003;8:15–19. - PubMed
-
- Munekage Y, et al. Cyclic electron flow around photosystem I is essential for photosynthesis. Nature. 2004;429:579–582. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
