

N-terminal truncations on L1 proteins of human papillomaviruses promote their soluble expression in Escherichia coli and self-assembly in vitro
- PMID: 30254257
- PMCID: PMC6156512
- DOI: 10.1038/s41426-018-0158-2
N-terminal truncations on L1 proteins of human papillomaviruses promote their soluble expression in Escherichia coli and self-assembly in vitro
Abstract
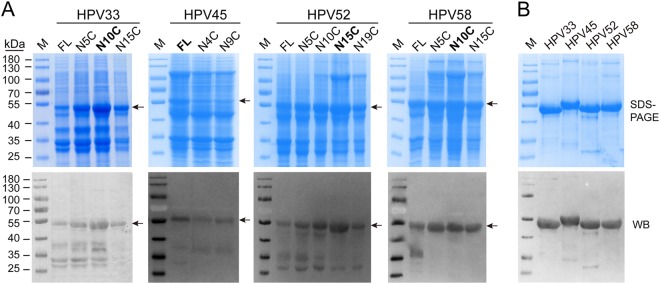
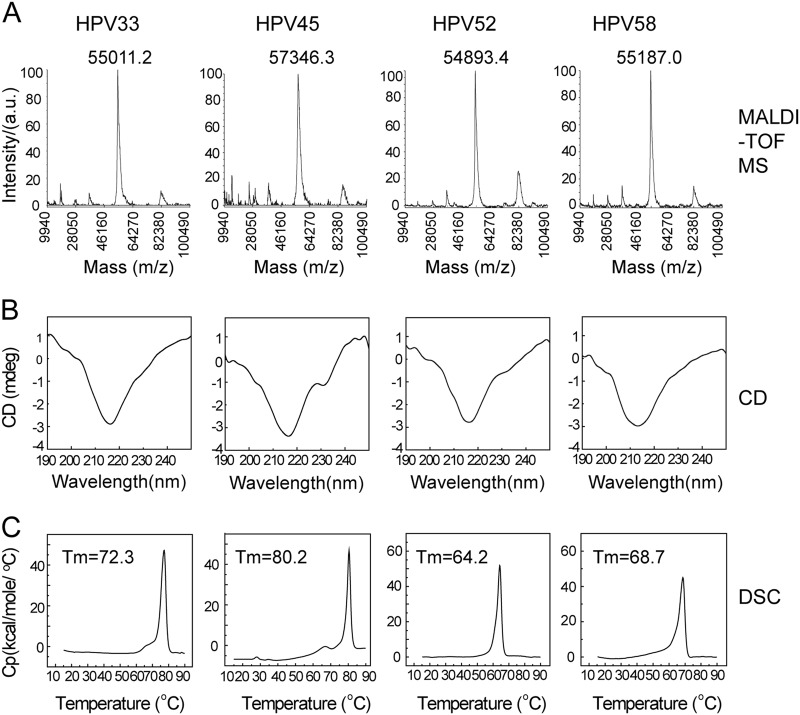
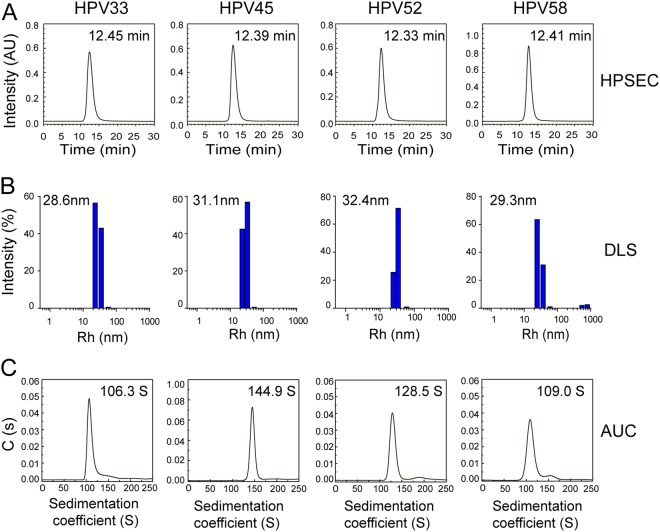
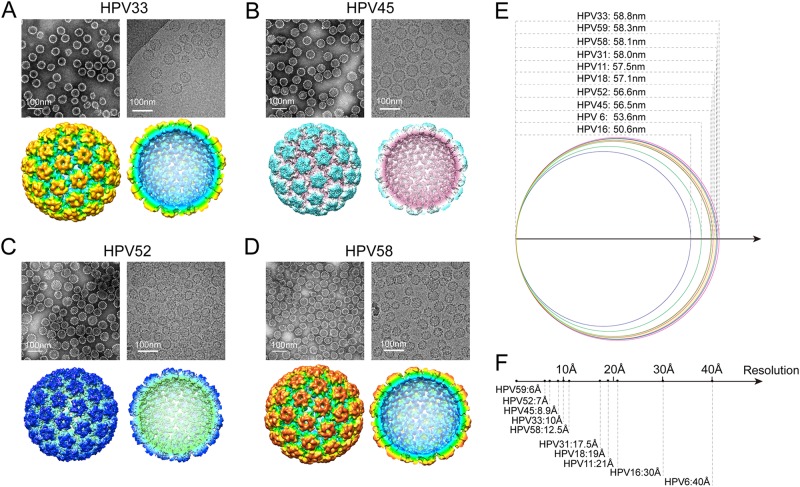
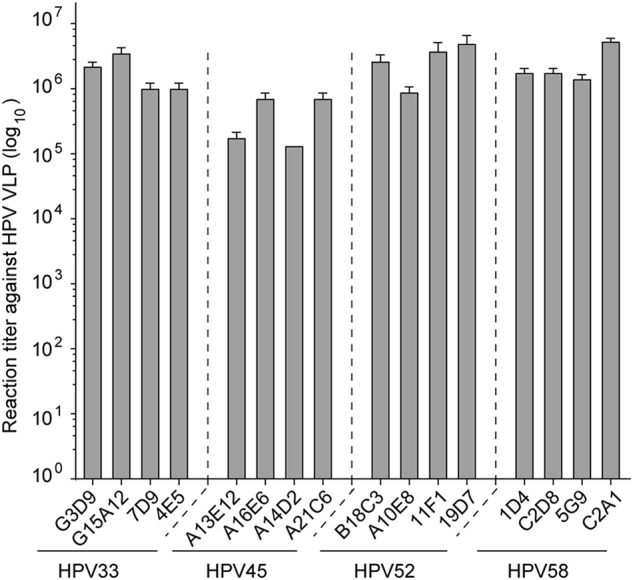
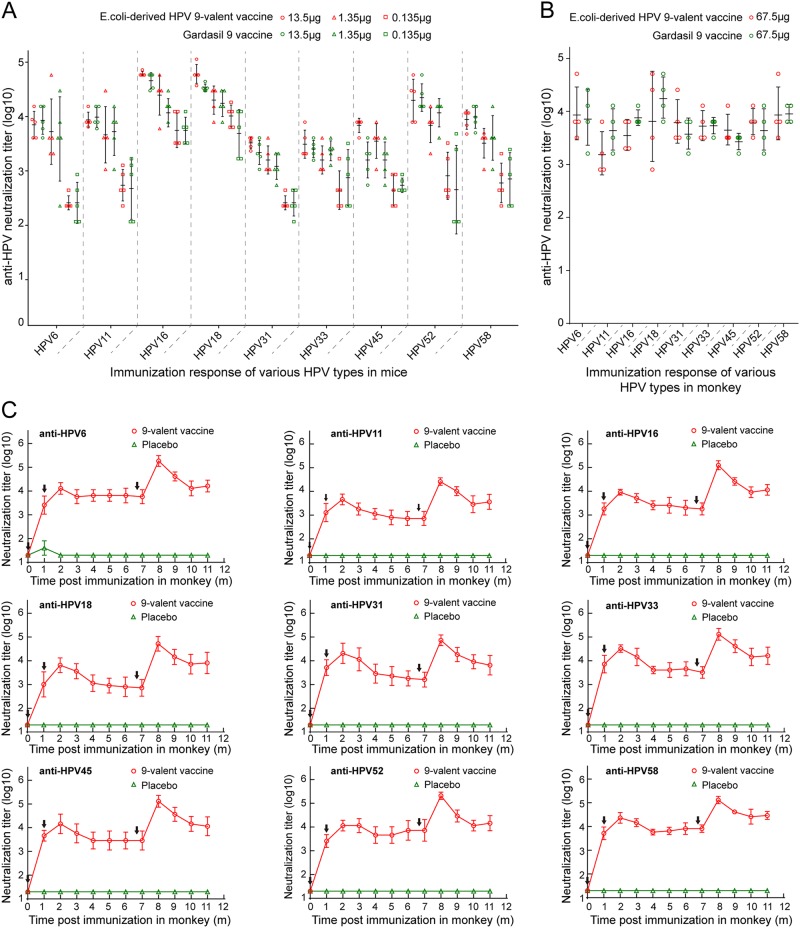
Human papillomavirus (HPV) is the causative agent in genital warts and nearly all cervical, anogenital, and oropharyngeal cancers. Nine HPV types (6, 11, 16, 18, 31, 33, 45, 52, and 58) are associated with about 90% of cervical cancers and 90% of genital warts. HPV neutralization by vaccine-elicited neutralizing antibodies can block viral infection and prevent HPV-associated diseases. However, there is only one commercially available HPV vaccine, Gardasil 9, produced from Saccharomyces cerevisiae that covers all nine types, raising the need for microbial production of broad-spectrum HPV vaccines. Here, we investigated whether N-terminal truncations of the major HPV capsid proteins L1, improve their soluble expression in Escherichia coli. We found that N-terminal truncations promoted the soluble expression of HPV 33 (truncated by 10 amino acids [aa]), 52 (15 aa), and 58 (10 aa). The resultant HPV L1 proteins were purified in pentamer form and extensively characterized with biochemical, biophysical, and immunochemical methods. The pentamers self-assembled into virus-like particles (VLPs) in vitro, and 3D cryo-EM reconstructions revealed that all formed T = 7 icosahedral particles having 50-60-nm diameters. Moreover, we formulated a nine-valent HPV vaccine candidate with aluminum adjuvant and L1 VLPs from four genotypes used in this study and five from previous work. Immunogenicity assays in mice and non-human primates indicated that this HPV nine-valent vaccine candidate elicits neutralizing antibody titers comparable to those induced by Gardasil 9. Our study provides a method for producing a nine-valent HPV vaccine in E. coli and may inform strategies for the soluble expression of other vaccine candidates.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Developments in L2-based human papillomavirus (HPV) vaccines.Virus Res. 2017 Mar 2;231:166-175. doi: 10.1016/j.virusres.2016.11.020. Epub 2016 Nov 23. Virus Res. 2017. PMID: 27889616 Free PMC article. Review.
-
Broad Cross-Protection Is Induced in Preclinical Models by a Human Papillomavirus Vaccine Composed of L1/L2 Chimeric Virus-Like Particles.J Virol. 2016 Jun 24;90(14):6314-25. doi: 10.1128/JVI.00449-16. Print 2016 Jul 15. J Virol. 2016. PMID: 27147749 Free PMC article.
-
Human papillomavirus L1 protein expressed in Escherichia coli self-assembles into virus-like particles that are highly immunogenic.Virus Res. 2016 Jul 15;220:97-103. doi: 10.1016/j.virusres.2016.04.017. Epub 2016 Apr 20. Virus Res. 2016. PMID: 27107614
-
Characterization of an Escherichia coli-derived human papillomavirus type 16 and 18 bivalent vaccine.Vaccine. 2017 Aug 16;35(35 Pt B):4637-4645. doi: 10.1016/j.vaccine.2017.06.084. Epub 2017 Jul 20. Vaccine. 2017. PMID: 28736197
-
Worldwide genetic variations in high-risk human papillomaviruses capsid L1 gene and their impact on vaccine efficiency.Gene. 2021 May 25;782:145533. doi: 10.1016/j.gene.2021.145533. Epub 2021 Feb 23. Gene. 2021. PMID: 33636291 Review.
Cited by
-
Recent Progress on the Versatility of Virus-Like Particles.Vaccines (Basel). 2020 Mar 20;8(1):139. doi: 10.3390/vaccines8010139. Vaccines (Basel). 2020. PMID: 32244935 Free PMC article. Review.
-
Engineering of Recombinant Human Papillomavirus 16 L1 Protein for Incorporation with para-Azido-L-Phenylalanine.J Microbiol Biotechnol. 2024 Sep 28;34(9):1926-1932. doi: 10.4014/jmb.2407.07033. Epub 2024 Aug 9. J Microbiol Biotechnol. 2024. PMID: 39155395 Free PMC article.
-
An in vivo gene amplification system for high level expression in Saccharomyces cerevisiae.Nat Commun. 2022 May 24;13(1):2895. doi: 10.1038/s41467-022-30529-8. Nat Commun. 2022. PMID: 35610221 Free PMC article.
-
Genetic diversity of HPV35 in Chad and the Central African Republic, two landlocked countries of Central Africa: A cross-sectional study.PLoS One. 2024 Jan 25;19(1):e0297054. doi: 10.1371/journal.pone.0297054. eCollection 2024. PLoS One. 2024. PMID: 38271382 Free PMC article.
-
High-Level Production of Soluble Cross-Reacting Material 197 in Escherichia coli Cytoplasm Due to Fine Tuning of the Target Gene's mRNA Structure.BioTech (Basel). 2023 Jan 11;12(1):9. doi: 10.3390/biotech12010009. BioTech (Basel). 2023. PMID: 36648835 Free PMC article.
References
-
- Munoz N, Castellsague X, de Gonzalez AB, Gissmann L. Chapter 1: HPV in the etiology of human cancer. Vaccine. 2006;24(Suppl 3):S3/1–S3/10. - PubMed
-
- Burchell AN, Winer RL, de Sanjose S, Franco EL. Chapter 6: Epidemiology and transmission dynamics of genital HPV infection. Vaccine. 2006;24(Suppl 3):S3/52–S3/561. - PubMed
MeSH terms
Substances
Grants and funding
- U1705283/National Natural Science Foundation of China (National Science Foundation of China)
- 31670935/National Natural Science Foundation of China (National Science Foundation of China)
- 81701637/National Natural Science Foundation of China (National Science Foundation of China)
- 2017J07005/Natural Science Foundation of Fujian Province (Fujian Provincial Natural Science Foundation)
LinkOut - more resources
Full Text Sources
Other Literature Sources