

Microenvironmental niche divergence shapes BRCA1-dysregulated ovarian cancer morphological plasticity
- PMID: 30254278
- PMCID: PMC6156340
- DOI: 10.1038/s41467-018-06130-3
Microenvironmental niche divergence shapes BRCA1-dysregulated ovarian cancer morphological plasticity
Abstract
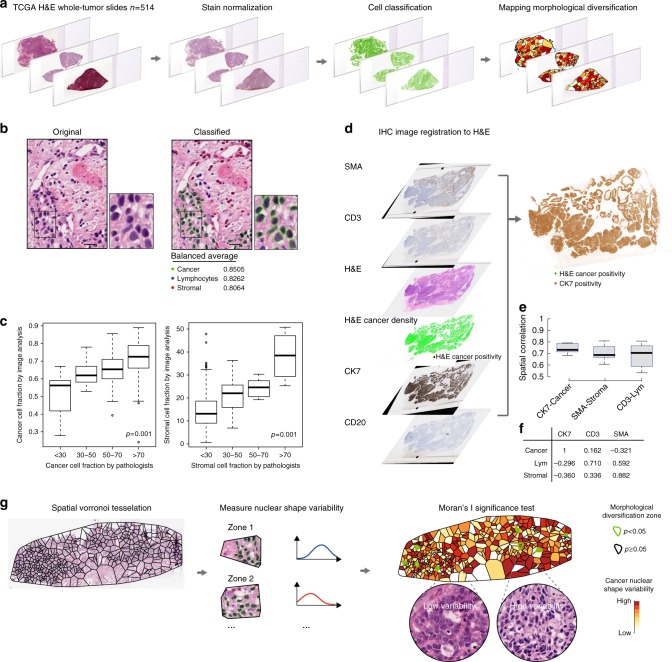
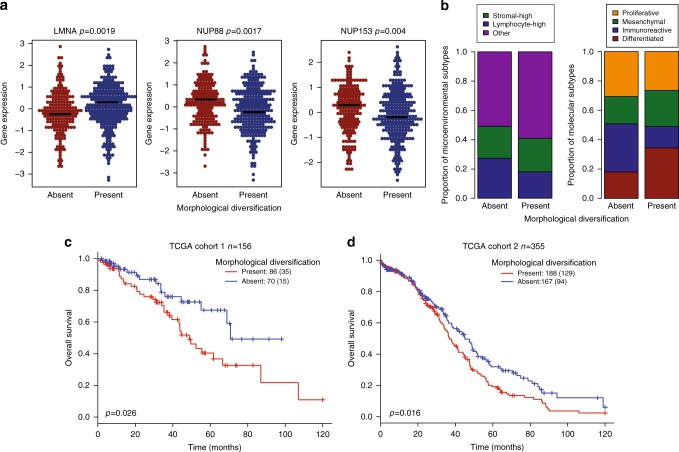
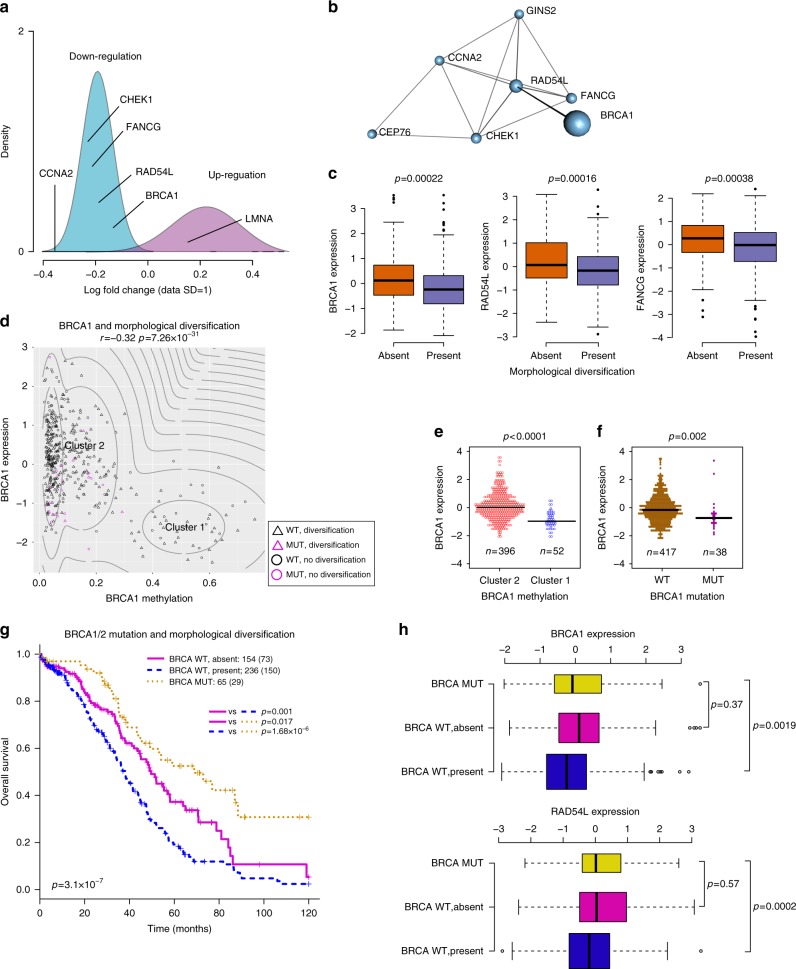
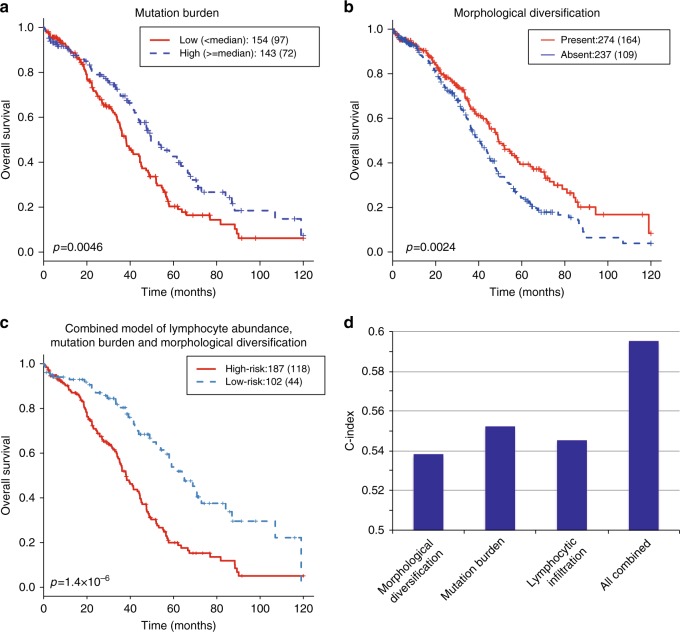
How tumor microenvironmental forces shape plasticity of cancer cell morphology is poorly understood. Here, we conduct automated histology image and spatial statistical analyses in 514 high grade serous ovarian samples to define cancer morphological diversification within the spatial context of the microenvironment. Tumor spatial zones, where cancer cell nuclei diversify in shape, are mapped in each tumor. Integration of this spatially explicit analysis with omics and clinical data reveals a relationship between morphological diversification and the dysregulation of DNA repair, loss of nuclear integrity, and increased disease mortality. Within the Immunoreactive subtype, spatial analysis further reveals significantly lower lymphocytic infiltration within diversified zones compared with other tumor zones, suggesting that even immune-hot tumors contain cells capable of immune escape. Our findings support a model whereby a subpopulation of morphologically plastic cancer cells with dysregulated DNA repair promotes ovarian cancer progression through positive selection by immune evasion.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
