

Differential Regulation of Adhesion and Phagocytosis of Resting and Activated Microglia by Dopamine
- PMID: 30254570
- PMCID: PMC6141656
- DOI: 10.3389/fncel.2018.00309
Differential Regulation of Adhesion and Phagocytosis of Resting and Activated Microglia by Dopamine
Abstract
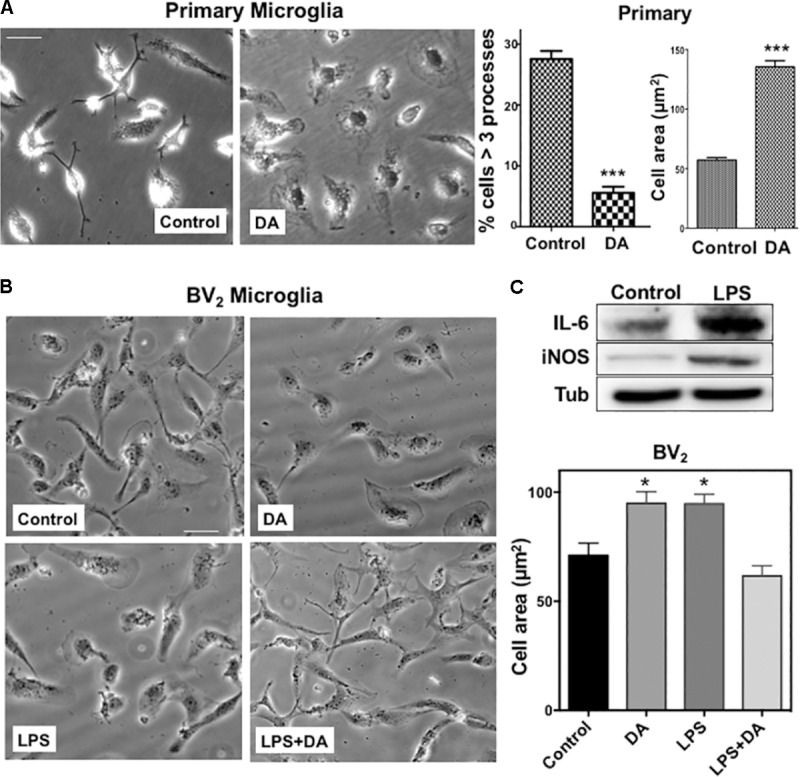
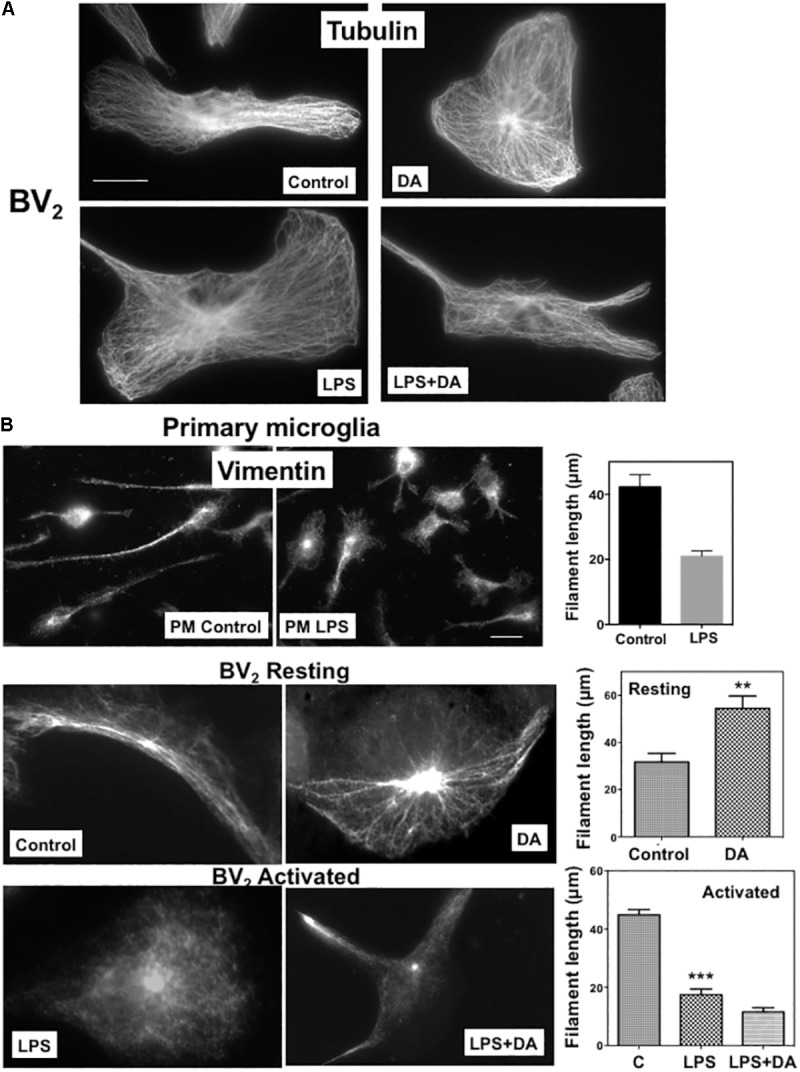
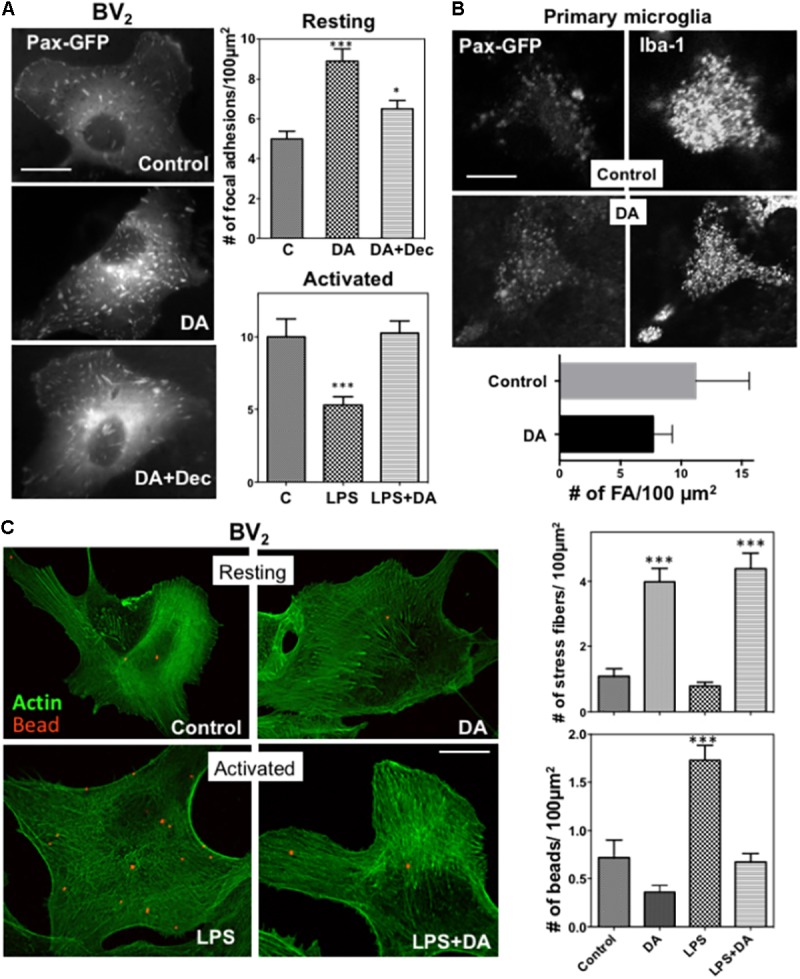
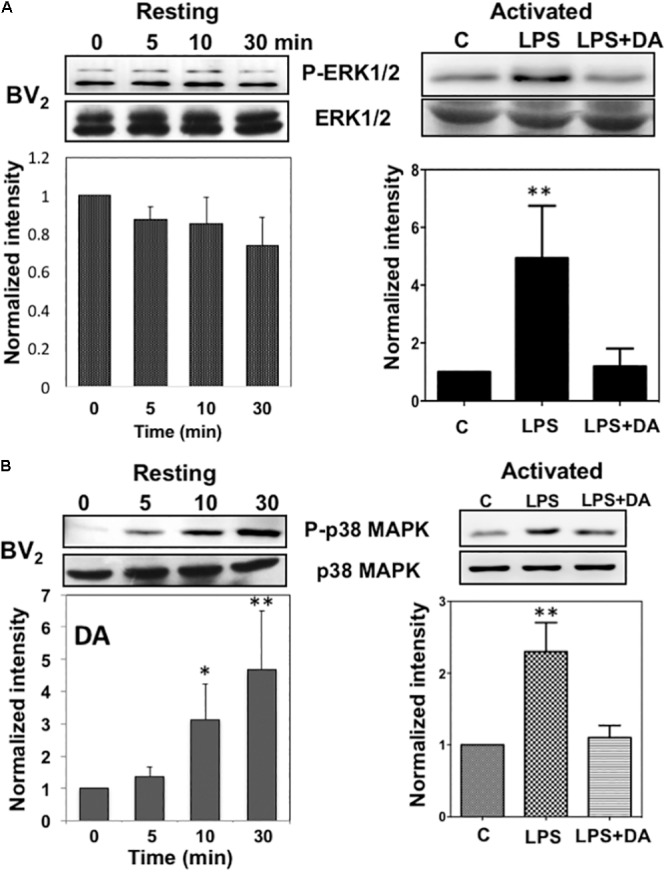
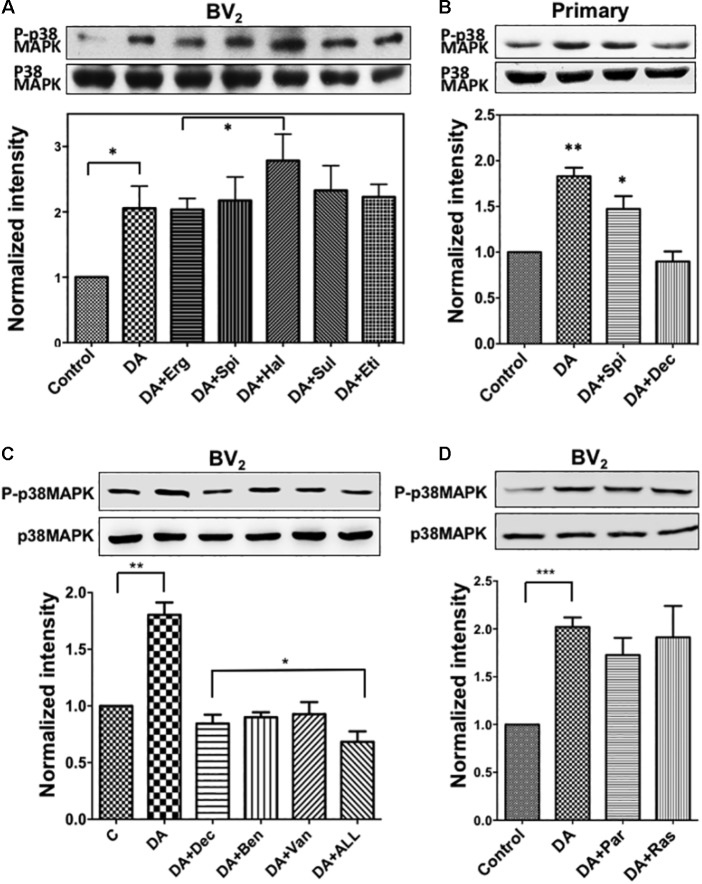
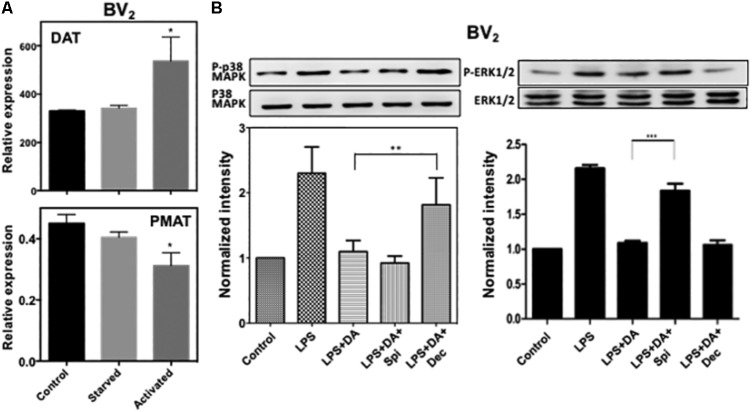
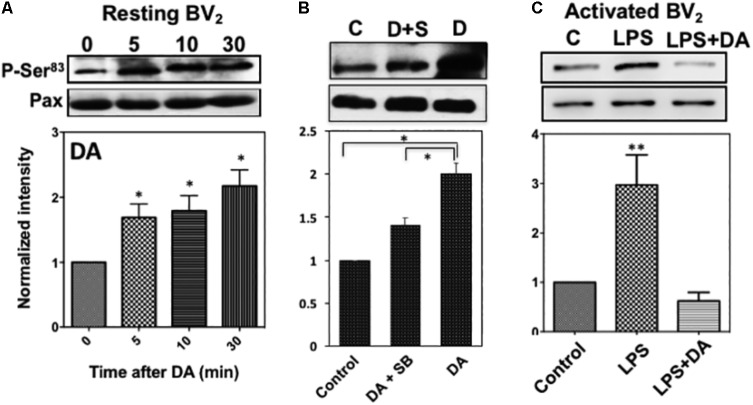
Microglia, the immune competent cells of the central nervous system (CNS), normally exist in a resting state characterized by a ramified morphology with many processes, and become activated to amoeboid morphology in response to brain injury, infection, and a variety of neuroinflammatory stimuli. Many studies focused on how neurotransmitters affect microglia activation in pathophysiological circumstances. In this study, we tried to gain mechanistic insights on how dopamine (DA) released from neurons modulates cellular functions of resting and activated microglia. DA induced the reduction of the number of cellular processes, the increase of cell adhesion/spreading, and the increase of vimentin filaments in resting primary and BV2 microglia. In contrast to resting cells, DA downregulated the cell spreading and phagocytosis of microglia activated by LPS. DA also significantly downregulated ERK1/2 phosphorylation in activated microglia, but not in resting microglia. Downregulation of ERK1/2 by DA in activated microglia required receptor signaling. In contrast, we found a significant increase of p38MAPK activity by DA treatment in resting, but not in activated microglia. These latter effects required the uptake of DA through the high-affinity transporter but did not require receptor signaling. Activation of p38MAPK resulted in the increase of focal adhesion number via phosphorylation of paxillin at Ser83. These results indicate that DA might have a differential, depending upon the activation stage of microglia, impact on cellular functions such as adhesion and phagocytosis.
Keywords: dopamine; microglia; p38MAPK; paxillin; phagocytosis.
Figures
Similar articles
-
A systems pharmacology-based approach to identify novel Kv1.3 channel-dependent mechanisms in microglial activation.J Neuroinflammation. 2017 Jun 26;14(1):128. doi: 10.1186/s12974-017-0906-6. J Neuroinflammation. 2017. PMID: 28651603 Free PMC article.
-
In Vitro Priming and Hyper-Activation of Brain Microglia: an Assessment of Phenotypes.Mol Neurobiol. 2019 Sep;56(9):6409-6425. doi: 10.1007/s12035-019-1529-y. Epub 2019 Feb 25. Mol Neurobiol. 2019. PMID: 30805836
-
Umbilical cord-derived mesenchymal stromal cells immunomodulate and restore actin dynamics and phagocytosis of LPS-activated microglia via PI3K/Akt/Rho GTPase pathway.Cell Death Discov. 2021 Mar 15;7(1):46. doi: 10.1038/s41420-021-00436-w. Cell Death Discov. 2021. PMID: 33723246 Free PMC article.
-
Phagocytic properties of microglia in vitro: implications for a role in multiple sclerosis and EAE.Microsc Res Tech. 2001 Jul 15;54(2):81-94. doi: 10.1002/jemt.1123. Microsc Res Tech. 2001. PMID: 11455615 Review.
-
Microglia: a sensor to threats in the nervous system?Res Virol. 1996 Mar-Jun;147(2-3):79-88. doi: 10.1016/0923-2516(96)80220-2. Res Virol. 1996. PMID: 8901425 Review.
Cited by
-
An integrated cytokine and kynurenine network as the basis of neuroimmune communication.Front Neurosci. 2022 Nov 24;16:1002004. doi: 10.3389/fnins.2022.1002004. eCollection 2022. Front Neurosci. 2022. PMID: 36507331 Free PMC article. Review.
-
Organic Cation Transporter (OCT/OCTN) Expression at Brain Barrier Sites: Focus on CNS Drug Delivery.Handb Exp Pharmacol. 2021;266:301-328. doi: 10.1007/164_2021_448. Handb Exp Pharmacol. 2021. PMID: 33674914 Free PMC article.
-
The impact of substance abuse on HIV-mediated neuropathogenesis in the current ART era.Brain Res. 2019 Dec 1;1724:146426. doi: 10.1016/j.brainres.2019.146426. Epub 2019 Aug 29. Brain Res. 2019. PMID: 31473221 Free PMC article. Review.
-
Molecular mechanisms underlying microglial sensing and phagocytosis in synaptic pruning.Neural Regen Res. 2024 Jun 1;19(6):1284-1290. doi: 10.4103/1673-5374.385854. Epub 2023 Sep 22. Neural Regen Res. 2024. PMID: 37905877 Free PMC article.
-
Apomorphine Reduces A53T α-Synuclein-Induced Microglial Reactivity Through Activation of NRF2 Signalling Pathway.Cell Mol Neurobiol. 2022 Nov;42(8):2673-2695. doi: 10.1007/s10571-021-01131-1. Epub 2021 Aug 20. Cell Mol Neurobiol. 2022. PMID: 34415465 Free PMC article.
References
-
- Bottalico B., Larsson I., Brodszki J., Hernandez-Andrade E., Casslén B., Marsál K., et al. (2004). Norepinephrine transporter (NET), serotonin transporter (SERT), vesicular monoamine transporter (VMAT2) and organic cation transporters (OCT1, 2 and EMT) in human placenta from pre-eclamptic and normotensive pregnancies. Placenta 25 518–529. 10.1016/j.placenta.2003.10.017 - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous