

Feasibility Theory Reconciles and Informs Alternative Approaches to Neuromuscular Control
- PMID: 30254579
- PMCID: PMC6141757
- DOI: 10.3389/fncom.2018.00062
Feasibility Theory Reconciles and Informs Alternative Approaches to Neuromuscular Control
Abstract
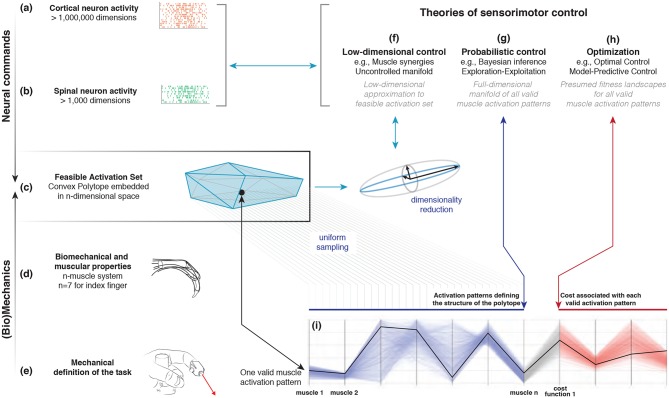
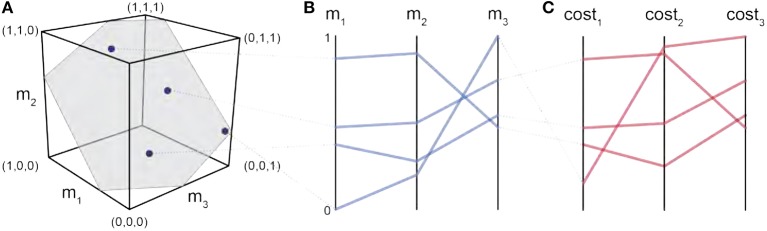
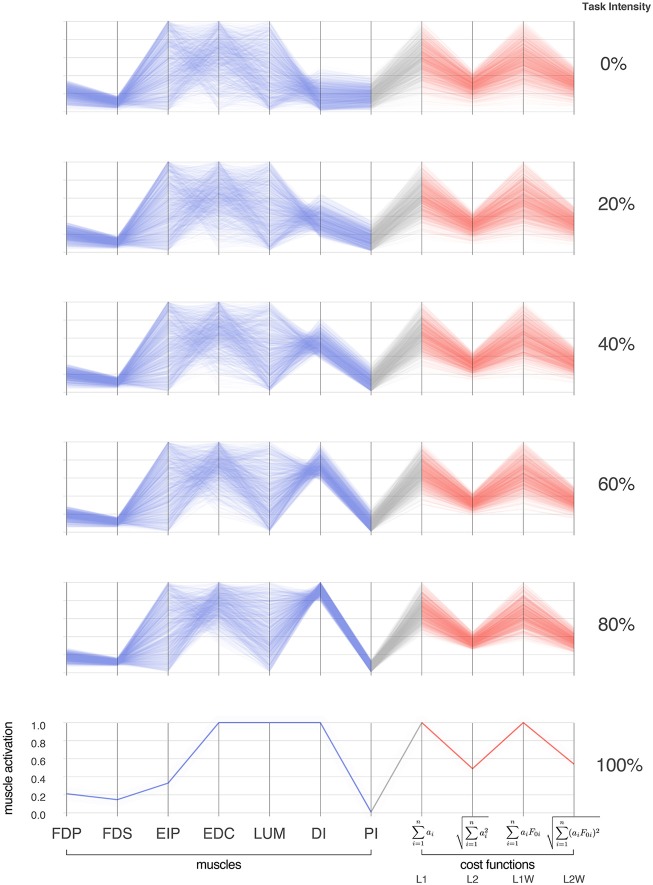
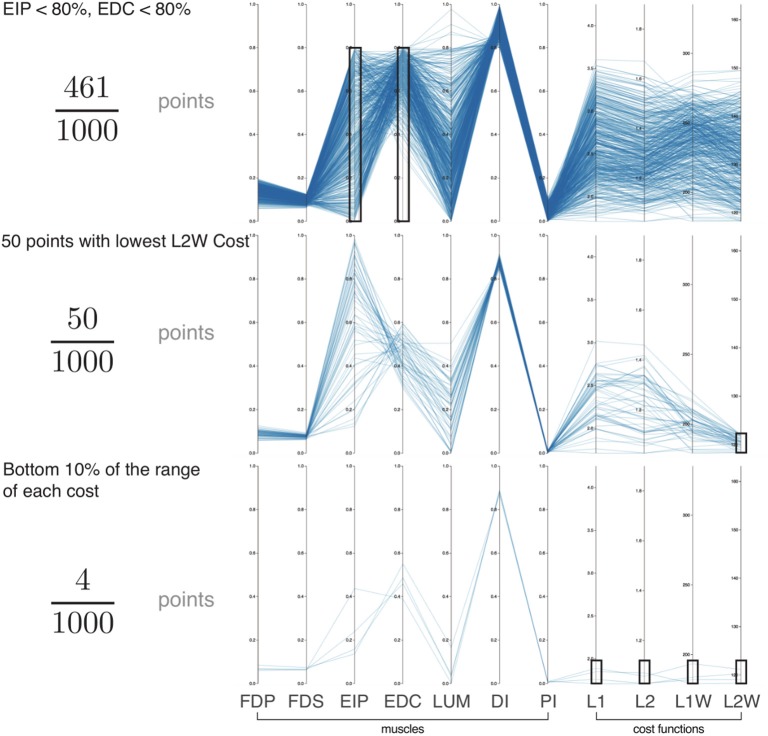
We present Feasibility Theory, a conceptual and computational framework to unify today's theories of neuromuscular control. We begin by describing how the musculoskeletal anatomy of the limb, the need to control individual tendons, and the physics of a motor task uniquely specify the family of all valid muscle activations that accomplish it (its 'feasible activation space'). For our example of producing static force with a finger driven by seven muscles, computational geometry characterizes-in a complete way-the structure of feasible activation spaces as 3-dimensional polytopes embedded in 7-D. The feasible activation space for a given task is the landscape where all neuromuscular learning, control, and performance must occur. This approach unifies current theories of neuromuscular control because the structure of feasible activation spaces can be separately approximated as either low-dimensional basis functions (synergies), high-dimensional joint probability distributions (Bayesian priors), or fitness landscapes (to optimize cost functions).
Keywords: dimensionality; feasibility; forces; motor control; neuromechanics; optimization; synergies; tendon-driven.
Figures
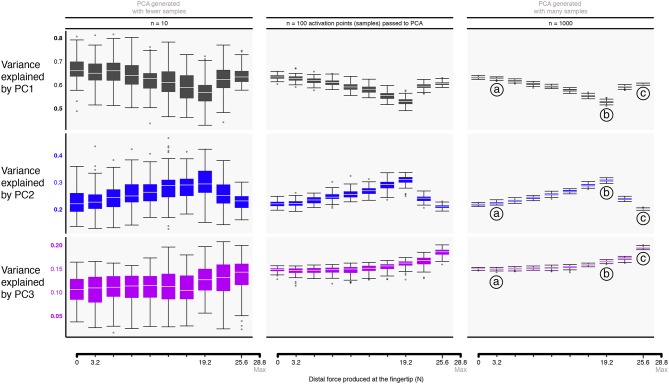
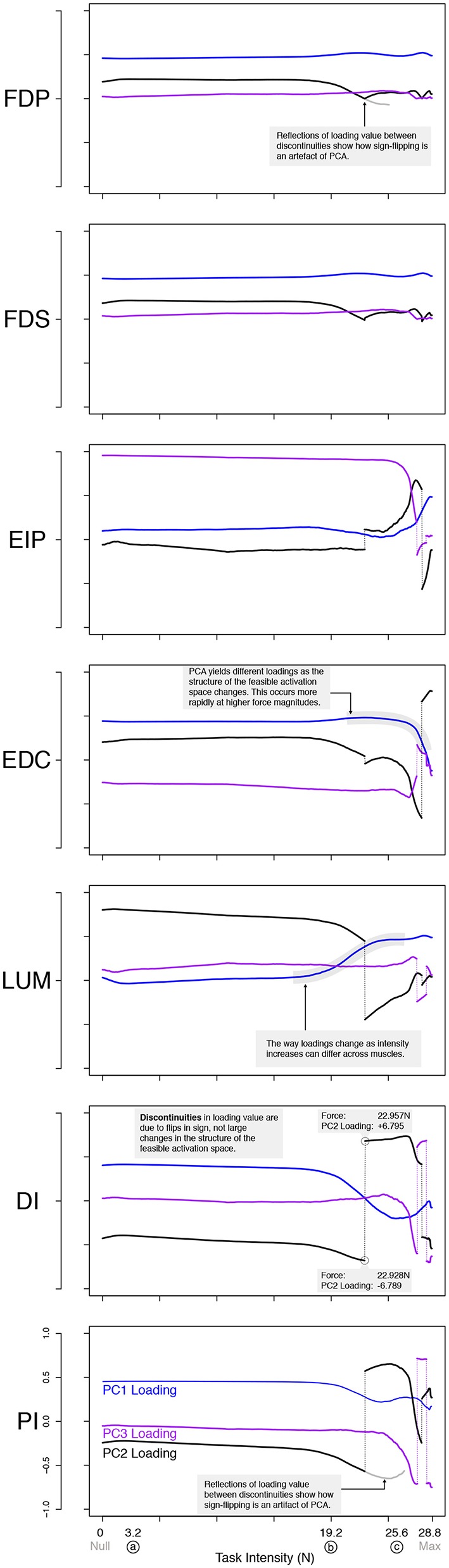
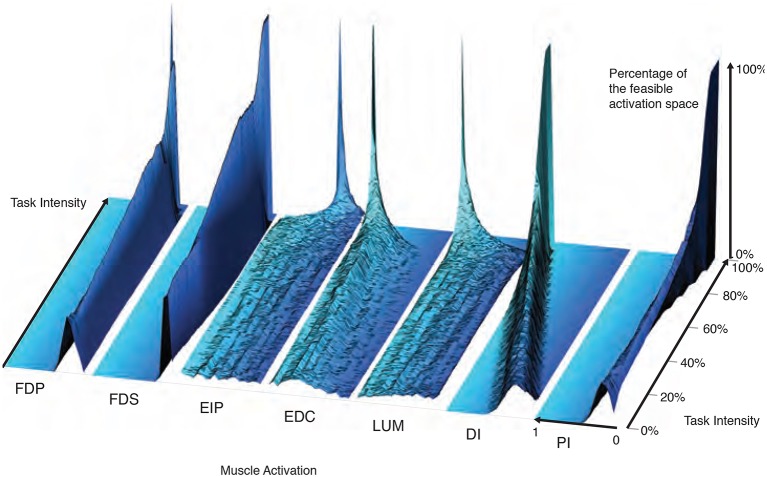
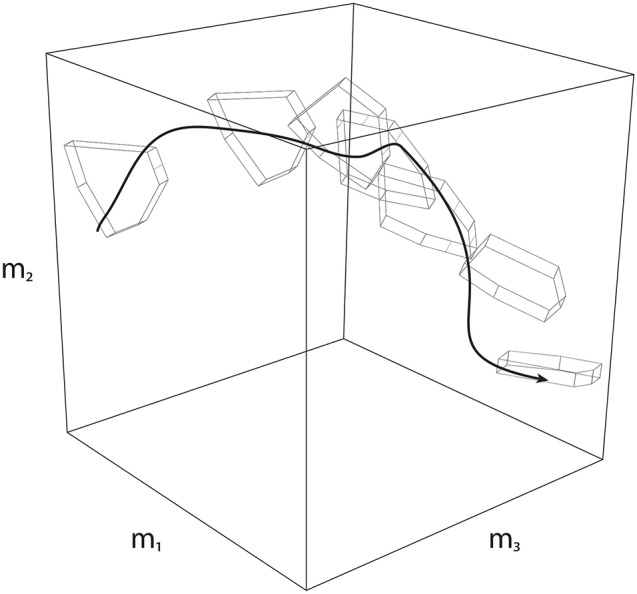
References
-
- Avis D., Fukuda K. (1992). A pivoting algorithm for convex hulls and vertex enumeration of arrangements and polyhedra. Dis. Comput. Geometry 8, 295–313. 10.1007/BF02293050 - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources