

Review
doi: 10.3389/fmicb.2018.02178.
eCollection 2018.
Adenoviromics: Mining the Human Adenovirus Species D Genome
Affiliations
- PMID: 30254627
- PMCID: PMC6141750
- DOI: 10.3389/fmicb.2018.02178
Item in Clipboard
Review
Adenoviromics: Mining the Human Adenovirus Species D Genome
Front Microbiol.
.
Erratum in
-
Corrigendum: Adenoviromics: Mining the Human Adenovirus Species D Genome.Front Microbiol. 2018 Dec 4;9:3005. doi: 10.3389/fmicb.2018.03005. eCollection 2018. Front Microbiol. 2018. PMID: 30555456 Free PMC article.
Abstract
Human adenovirus (HAdV) infections cause disease world-wide. Whole genome sequencing has now distinguished 90 distinct genotypes in 7 species (A-G). Over half of these 90 HAdVs fall within species D, with essentially all of the HAdV-D whole genome sequences generated in the last decade. Herein, we describe recent new findings made possible by mining of this expanded genome database, and propose future directions to elucidate new functional elements and new functions for previously known viral components.
Keywords: adenovirus; evolution; genome; interactome; transcription factor.
Figures
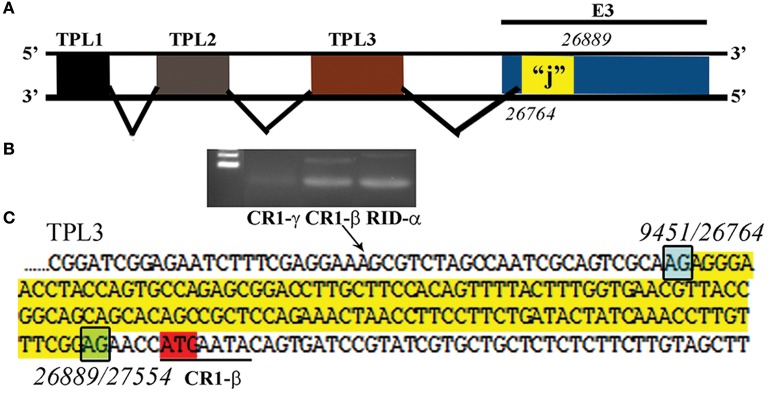
Putative “j”-leader located within the CR1-α E3 gene. (A) Schematic for the location of a newly detected leader (“j”-leader) embedded within the E3 CRI-α gene, experimentally determined to be spliced to some, but not all mRNAs of the E3 genes. (B) Gel photomicrograph of mRNA transcripts amplified with forward primer from TPL1 and reverse primers from CR1-γ, CR1-β, and RID-α. Primers were chosen to elicit similarly sized bands to facilitate subsequent sequencing. (C) Nucleotide sequence of the PCR product for CR1-β. The putative j-leader sequence and splice sites are shown in yellow and green, respectively. Note an additional 4 nucleotide 5′UTR (AACC) prior to the CR1-β start site (red). The 5′UTR in (C) prior to the splice site for the j-leader is from TPL3. Adapted from Ramke et al. (2017) with permission.
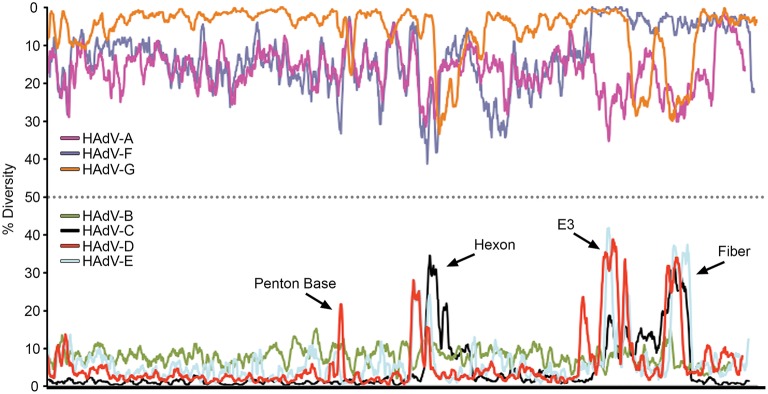
Nucleotide diversity plots, by HAdV species, generated with DnaSP, represent the average number of nucleotide differences per site between each type in every HAdV species. The % diversity is calculated on the y-axis; the x-axis illustrates the nucleotide position on the genome. HAdV-Ds (red line on bottom half of plot) show particular diversity in the penton base, hexon, E3, and fiber coding regions, with otherwise very high conservation. From Robinson et al. (2013a) with permission.
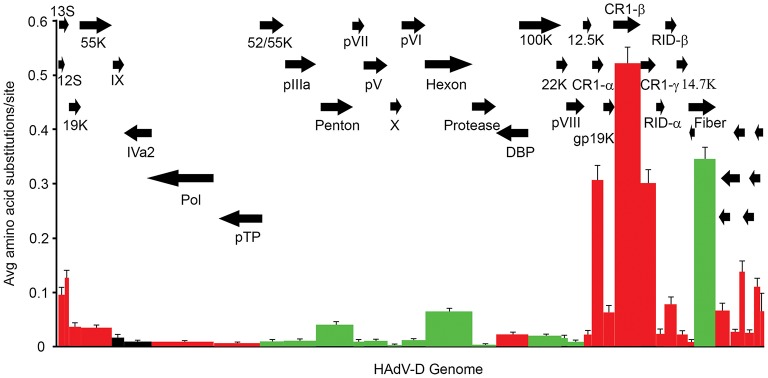
Amino acid diversity calculated in MEGA 4.02, measuring the average amino acid substitution for each HAdV-D protein. Each bar in the graph corresponds to a protein as represented by arrows. Red = early genes. Dark green = late genes. Black = intermediate genes. The hypervariable loops of penton base and hexon proteins were also analyzed separately (light green) and showed particularly high levels of amino acid substitutions. From Robinson et al. (2013a) with permission.
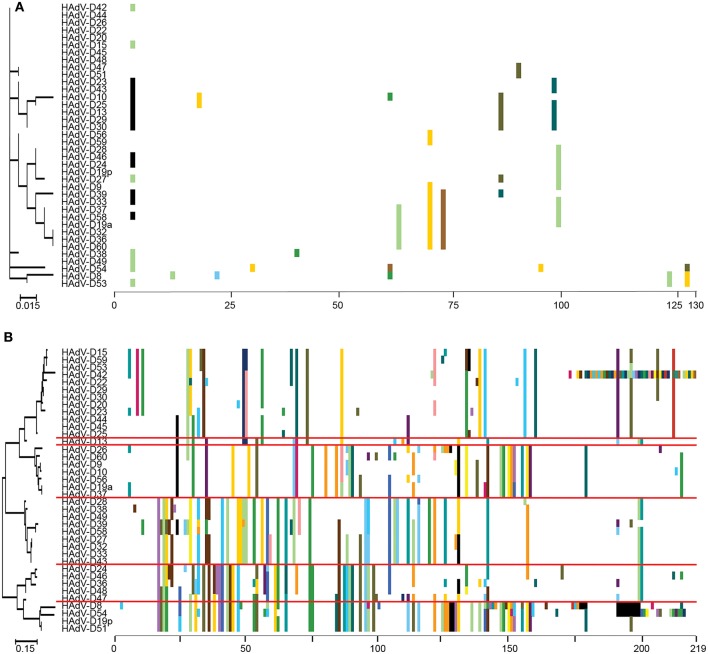
Proteotyping analysis comparing the HAdV-D E3 14.7K (A) and CR1α (B) proteins. The 14.7K protein was conserved, while CR1α demonstrated 6 unique proteotypes. Maximum likelihood phylogenetic trees are shown to the left for each putative protein, and amino acid signatures to the right. The scale bar at the bottom left of each sub-figure denotes the phylogenetic distance reflected in horizontal dimension of the respective tree. To construct the amino acid signatures shown, each amino acid was assigned a unique color (upper right corner), consensus amino acids at each position across all 38 viruses were assigned white, and gaps in the alignment were colored black. Horizontal red lines delineate distinct proteotypes. Adapted from. Singh et al. (2013) with permission.
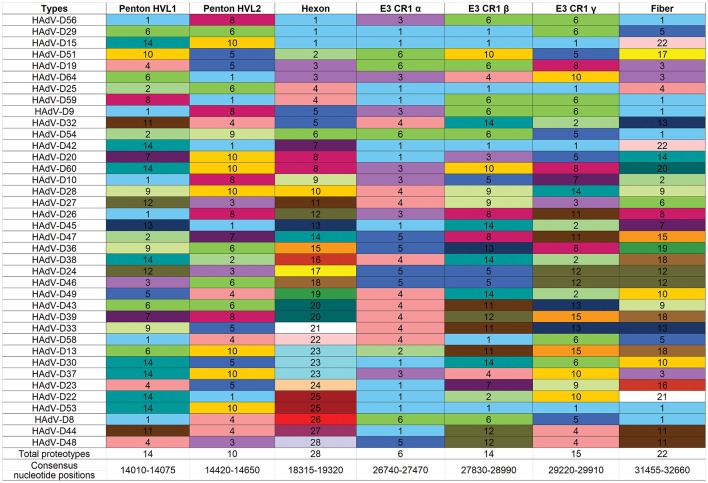
Proteotyping for 38 HAdV-Ds, sorted for the hexon proteotype column. Numbers and colors are arbitrary, and distinguish distinct proteotypes. Recombinants can be identified by rows. For example, HadV-D56, -D29, and -D15 fall within the same proteotype and are predicted to share highly similar nucleotide sequences for their respective hexon hypervariable regions (as confirmed by Singh et al., 2015). For HAdV-D29 and –D15, the recombination event extended through the E3 CR1β ORF gene and then ended.
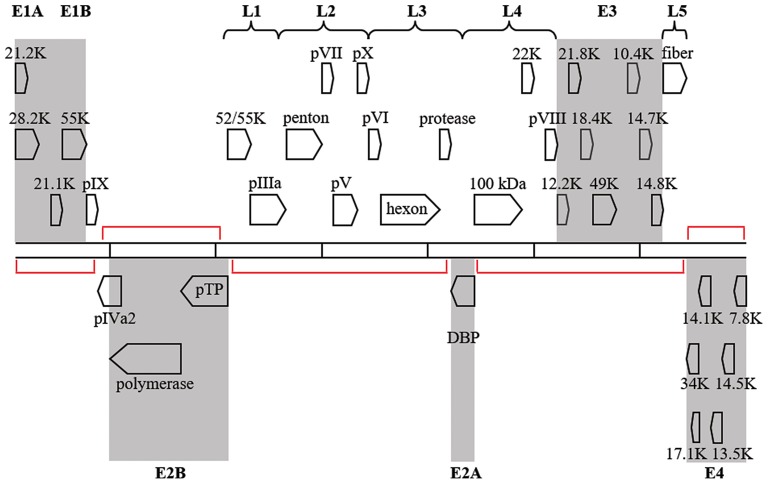
Transcription map for HAdV-D37. Genes are divided by early (shaded) or late expression. Red brackets denote large areas of “noncoding” DNA, but many additional, smaller, potential coding regions exist between and within known genes. Adapted from Robinson et al. (2008) with permission.
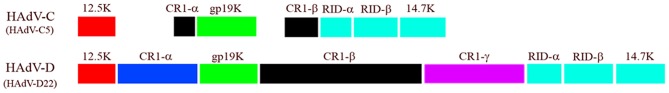
Comparison of E3 transcription unit from HAdV-C and -D. Note in particular the difference in ORF size between CR1β of the two HAdV species. Adapted from Robinson et al. (2011c) with permission.
References
-
- Andersson M., McMichael A., Peterson P. A. (1987). Reduced allorecognition of adenovirus-2 infected cells. J. Immunol. 138, 3960–3966. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources