

Cyclin-dependent kinase modulates budding yeast Rad5 stability during cell cycle
- PMID: 30256854
- PMCID: PMC6157869
- DOI: 10.1371/journal.pone.0204680
Cyclin-dependent kinase modulates budding yeast Rad5 stability during cell cycle
Abstract
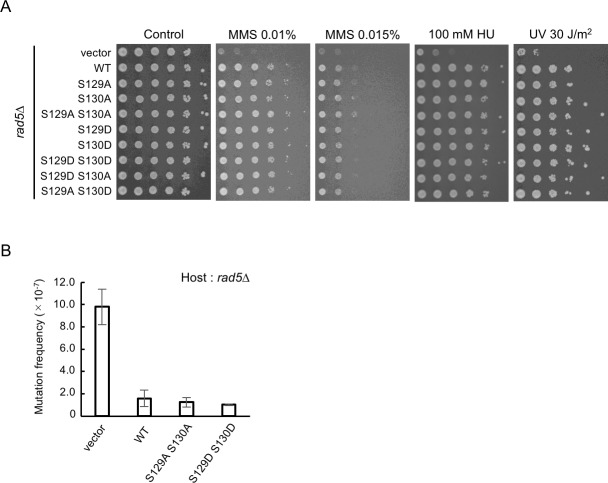
The DNA damage tolerance (DDT) pathway facilitates the bypass of the fork-blocking lesions without removing them through either translesion DNA synthesis or error-free damage bypass mechanism. The Saccharomyces cerevisiae Rad5 is a multi-functional protein involved in the error-free branch of the DDT pathway, and its protein level periodically fluctuates through the cell cycle; however, the mechanistic basis and functional importance of the Rad5 level for the cell cycle regulation remain unclear. Here, we show that Rad5 is predominantly phosphorylated on serine 130 (S130) during S/G2 phase and that this modification depends on the cyclin-dependent kinase Cdc28/CDK1. We also show that the phosphorylated Rad5 species at S130 exhibit a relatively short half-life compared with non-phosphorylated Rad5 moiety, and that the Rad5 protein is partially stabilized in phosphorylation-defective rad5 S130A cells. Importantly, the elimination of this modification results in a defective cell-cycle dependent Rad5 oscillation pattern. Together, our results demonstrate that CDK1 modulates Rad5 stability by phosphorylation during the cell cycle, suggesting a crosstalk between the phosphorylation and degradation of Rad5.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




Similar articles
-
Rad5 coordinates translesion DNA synthesis pathway by recognizing specific DNA structures in saccharomyces cerevisiae.Curr Genet. 2018 Aug;64(4):889-899. doi: 10.1007/s00294-018-0807-y. Epub 2018 Feb 2. Curr Genet. 2018. PMID: 29396601
-
Yeast Rad5 protein required for postreplication repair has a DNA helicase activity specific for replication fork regression.Mol Cell. 2007 Oct 12;28(1):167-75. doi: 10.1016/j.molcel.2007.07.030. Mol Cell. 2007. PMID: 17936713 Free PMC article.
-
The Rad5 helicase activity is dispensable for error-free DNA post-replication repair.DNA Repair (Amst). 2014 Apr;16:74-83. doi: 10.1016/j.dnarep.2014.02.016. Epub 2014 Mar 13. DNA Repair (Amst). 2014. PMID: 24674630
-
Role of yeast Rad5 and its human orthologs, HLTF and SHPRH in DNA damage tolerance.DNA Repair (Amst). 2010 Mar 2;9(3):257-67. doi: 10.1016/j.dnarep.2009.12.013. Epub 2010 Jan 21. DNA Repair (Amst). 2010. PMID: 20096653 Review.
-
The DNA damage tolerance factor Rad5 and telomere replication.Curr Genet. 2025 May 26;71(1):11. doi: 10.1007/s00294-025-01315-y. Curr Genet. 2025. PMID: 40418329 Free PMC article. Review.
Cited by
-
Qingjie Fuzheng granules inhibit colorectal cancer cell growth by the PI3K/AKT and ERK pathways.World J Gastrointest Oncol. 2019 May 15;11(5):377-392. doi: 10.4251/wjgo.v11.i5.377. World J Gastrointest Oncol. 2019. PMID: 31139308 Free PMC article.
-
A Conserved Histone H3-H4 Interface Regulates DNA Damage Tolerance and Homologous Recombination during the Recovery from Replication Stress.Mol Cell Biol. 2021 Mar 24;41(4):e00044-20. doi: 10.1128/MCB.00044-20. Print 2021 Mar 24. Mol Cell Biol. 2021. PMID: 33526454 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
