

G protein-coupled receptors differentially regulate glycosylation and activity of the inwardly rectifying potassium channel Kir7.1
- PMID: 30257863
- PMCID: PMC6240878
- DOI: 10.1074/jbc.RA118.003238
G protein-coupled receptors differentially regulate glycosylation and activity of the inwardly rectifying potassium channel Kir7.1
Abstract
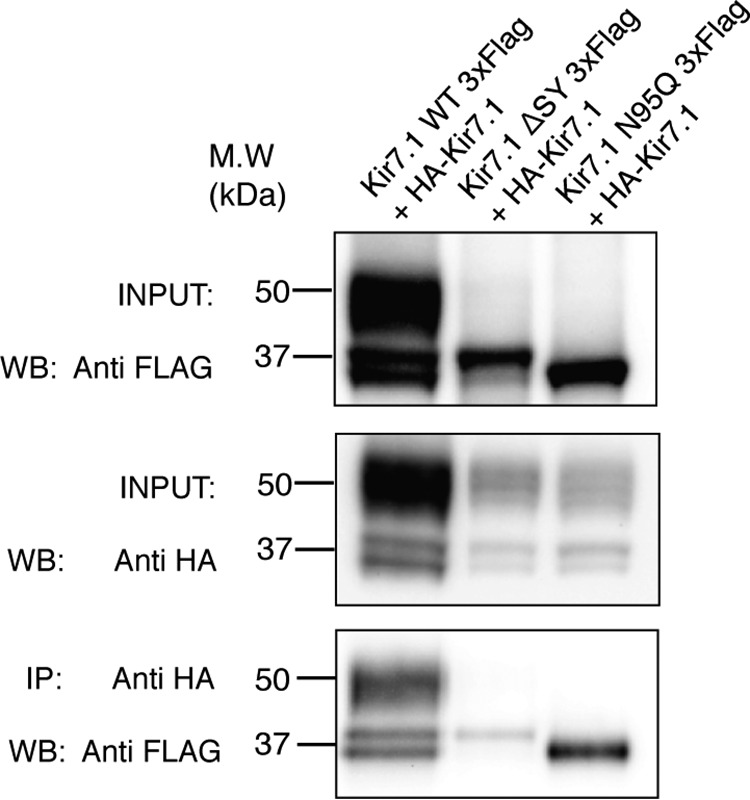
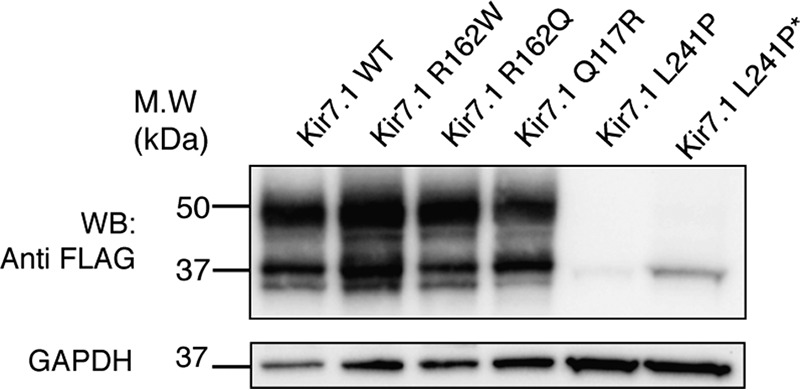
Kir7.1 is an inwardly rectifying potassium channel with important roles in the regulation of the membrane potential in retinal pigment epithelium, uterine smooth muscle, and hypothalamic neurons. Regulation of G protein-coupled inwardly rectifying potassium (GIRK) channels by G protein-coupled receptors (GPCRs) via the G protein βγ subunits has been well characterized. However, how Kir channels are regulated is incompletely understood. We report here that Kir7.1 is also regulated by GPCRs, but through a different mechanism. Using Western blotting analysis, we observed that multiple GPCRs tested caused a striking reduction in the complex glycosylation of Kir7.1. Further, GPCR-mediated reduction of Kir7.1 glycosylation in HEK293T cells did not alter its expression at the cell surface but decreased channel activity. Of note, mutagenesis of the sole Kir7.1 glycosylation site reduced conductance and open probability, as indicated by single-channel recording. Additionally, we report that the L241P mutation of Kir7.1 associated with Lebers congenital amaurosis (LCA), an inherited retinal degenerative disease, has significantly reduced complex glycosylation. Collectively, these results suggest that Kir7.1 channel glycosylation is essential for function, and this activity within cells is suppressed by most GPCRs. The melanocortin-4 receptor (MC4R), a GPCR previously reported to induce ligand-regulated activity of this channel, is the only GPCR tested that does not have this effect on Kir7.1.
Keywords: G protein–coupled receptor (GPCR); glycosylation; ion channel; potassium channel; signal transduction.
© 2018 Carrington et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures








References
-
- Nakamura N., Suzuki Y., Sakuta H., Ookata K., Kawahara K., and Hirose S. (1999) Inwardly rectifying K+ channel Kir7.1 is highly expressed in thyroid follicular cells, intestinal epithelial cells and choroid plexus epithelial cells: implication for a functional coupling with Na+,K+-ATPase. Biochem. J. 342, 329–336 10.1042/bj3420329, 10.1042/0264-6021:3420329 - DOI - PMC - PubMed
-
- Pattnaik B. R., Tokarz S., Asuma M. P., Schroeder T., Sharma A., Mitchell J. C., Edwards A. O., and Pillers D. A. (2013) Snowflake vitreoretinal degeneration (SVD) mutation R162W provides new insights into Kir7.1 ion channel structure and function. PLoS One 8, e71744 10.1371/journal.pone.0071744 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
