

PAX5-ELN oncoprotein promotes multistep B-cell acute lymphoblastic leukemia in mice
- PMID: 30257940
- PMCID: PMC6187160
- DOI: 10.1073/pnas.1721678115
PAX5-ELN oncoprotein promotes multistep B-cell acute lymphoblastic leukemia in mice
Abstract
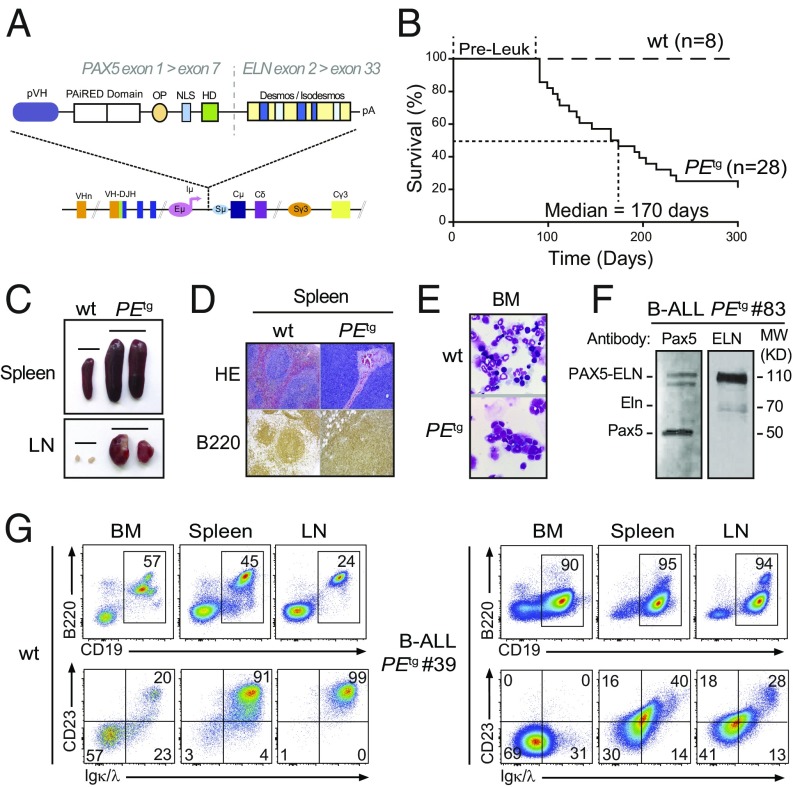
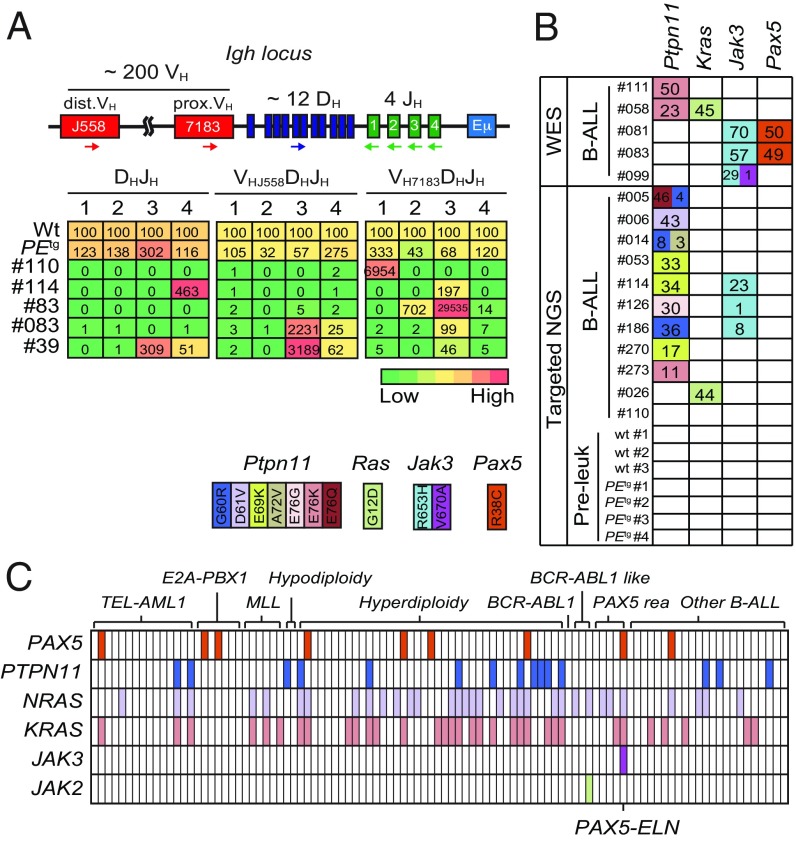
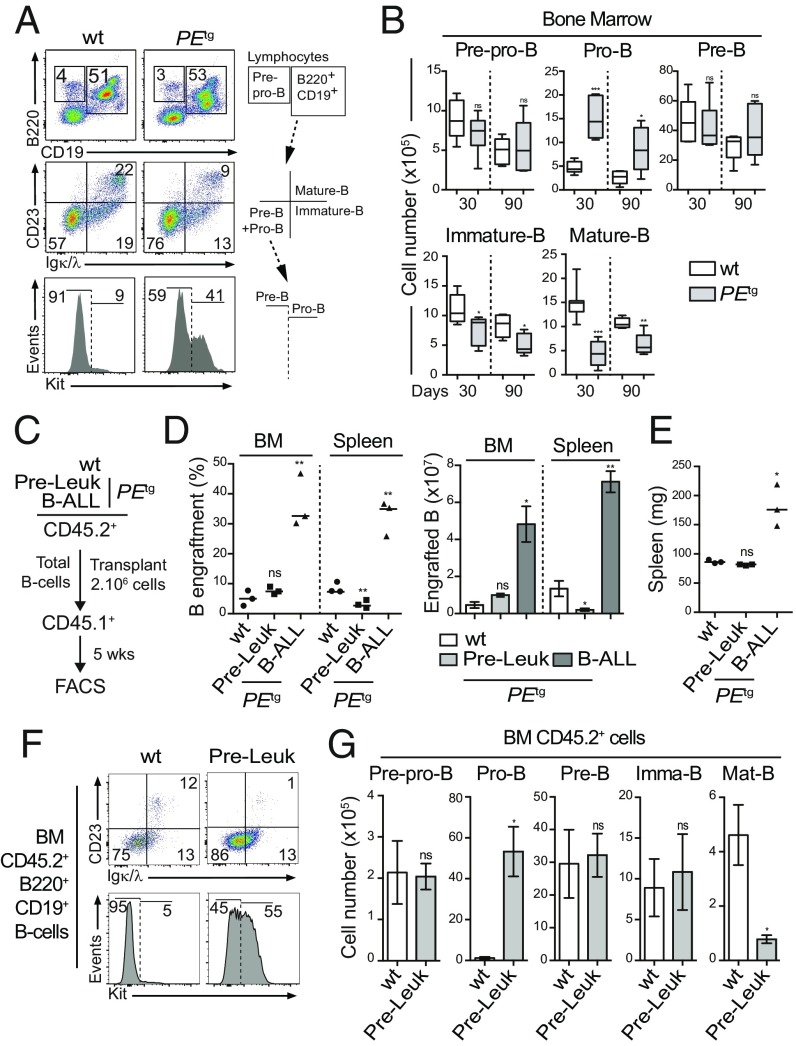
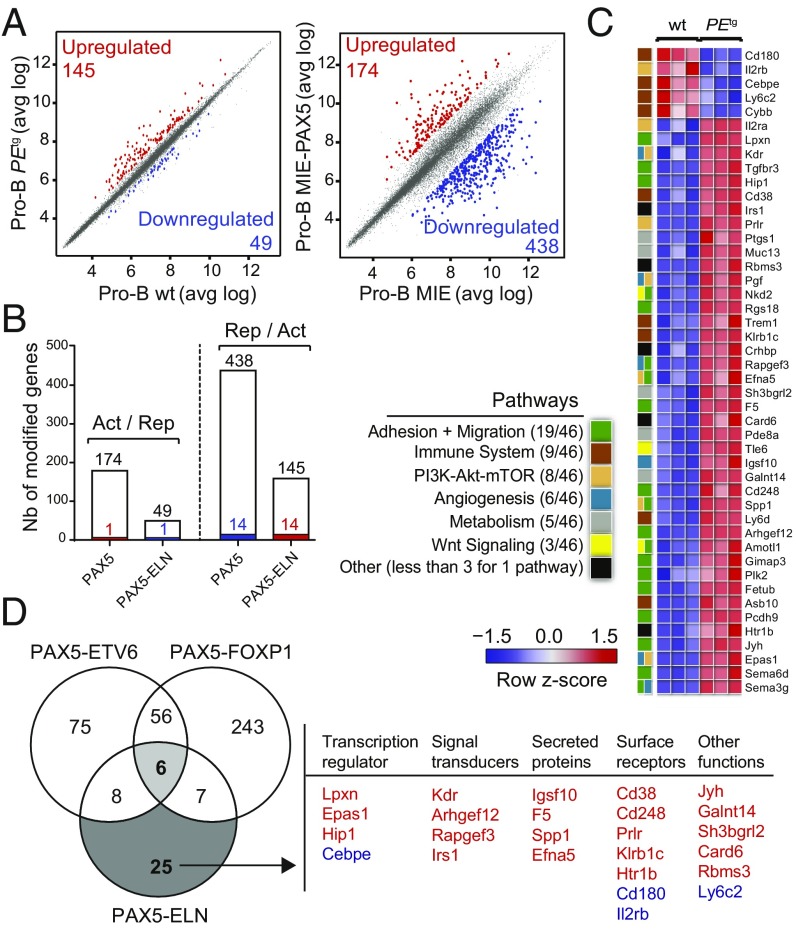
PAX5 is a well-known haploinsufficient tumor suppressor gene in human B-cell precursor acute lymphoblastic leukemia (B-ALL) and is involved in various chromosomal translocations that fuse a part of PAX5 with other partners. However, the role of PAX5 fusion proteins in B-ALL initiation and transformation is ill-known. We previously reported a new recurrent t(7;9)(q11;p13) chromosomal translocation in human B-ALL that juxtaposed PAX5 to the coding sequence of elastin (ELN). To study the function of the resulting PAX5-ELN fusion protein in B-ALL development, we generated a knockin mouse model in which the PAX5-ELN transgene is expressed specifically in B cells. PAX5-ELN-expressing mice efficiently developed B-ALL with an incidence of 80%. Leukemic transformation was associated with recurrent secondary mutations on Ptpn11, Kras, Pax5, and Jak3 genes affecting key signaling pathways required for cell proliferation. Our functional studies demonstrate that PAX5-ELN affected B-cell development in vitro and in vivo featuring an aberrant expansion of the pro-B cell compartment at the preleukemic stage. Finally, our molecular and computational approaches identified PAX5-ELN-regulated gene candidates that establish the molecular bases of the preleukemic state to drive B-ALL initiation. Hence, our study provides a new in vivo model of human B-ALL and strongly implicates PAX5 fusion proteins as potent oncoproteins in leukemia development.
Keywords: B-cell acute lymphoblastic leukemia; PAX5 fusion proteins; engineered mouse models; leukemia initiation; oncogenic transformation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Armstrong SA, Look AT. Molecular genetics of acute lymphoblastic leukemia. J Clin Oncol. 2005;23:6306–6315. - PubMed
-
- Hardy RR, et al. B-cell commitment, development and selection. Immunol Rev. 2000;175:23–32. - PubMed
-
- Adams B, et al. Pax-5 encodes the transcription factor BSAP and is expressed in B lymphocytes, the developing CNS, and adult testis. Genes Dev. 1992;6:1589–1607. - PubMed
-
- Urbánek P, Wang ZQ, Fetka I, Wagner EF, Busslinger M. Complete block of early B cell differentiation and altered patterning of the posterior midbrain in mice lacking Pax5/BSAP. Cell. 1994;79:901–912. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
