

Cell type-specific activation of mitogen-activated protein kinase in D1 receptor-expressing neurons of the nucleus accumbens potentiates stimulus-reward learning in mice
- PMID: 30258218
- PMCID: PMC6158283
- DOI: 10.1038/s41598-018-32840-1
Cell type-specific activation of mitogen-activated protein kinase in D1 receptor-expressing neurons of the nucleus accumbens potentiates stimulus-reward learning in mice
Abstract
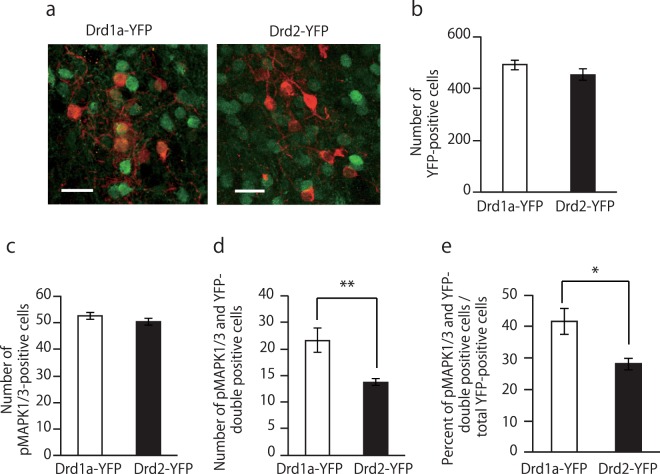
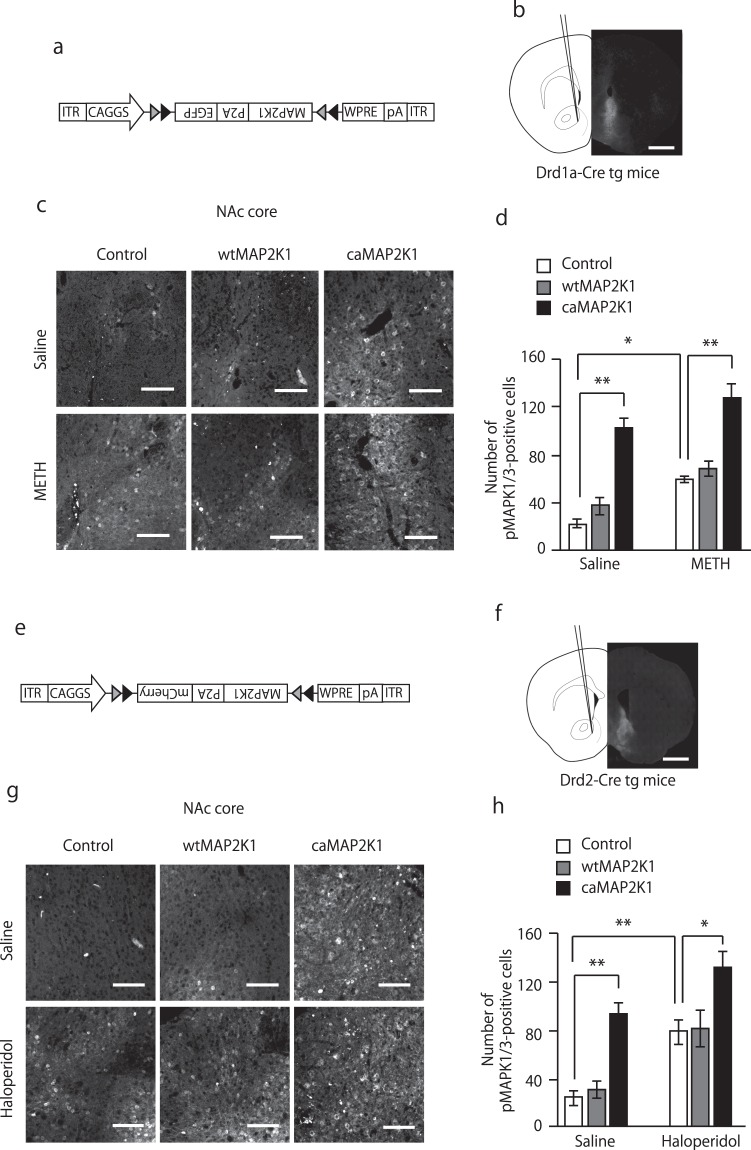
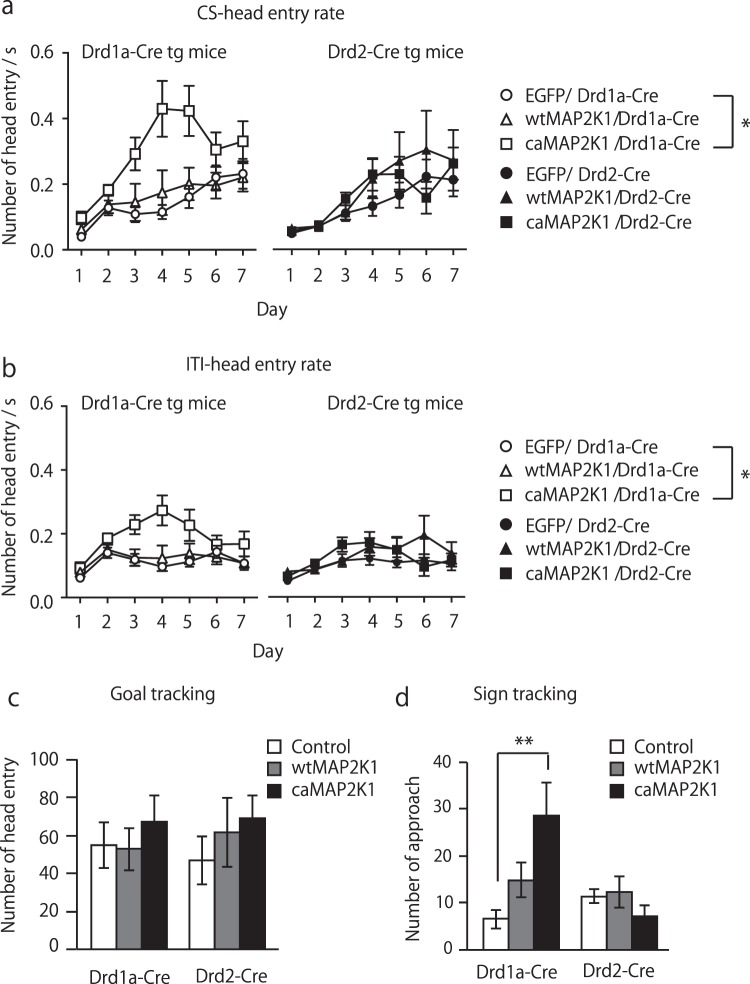
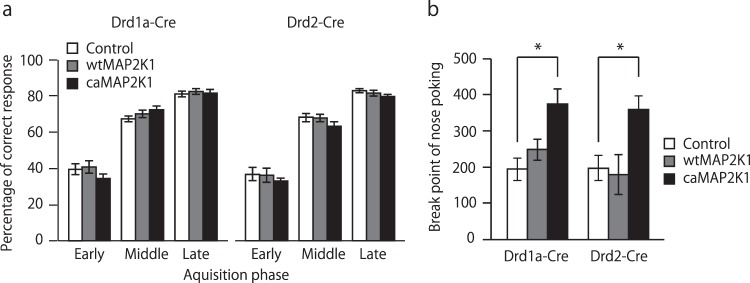
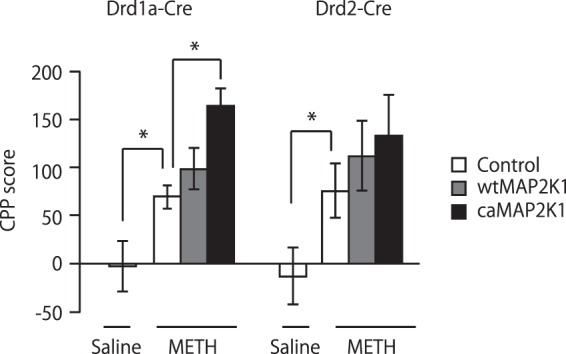
Medium spiny neurons (MSN) in the nucleus accumbens (NAc) are a fundamental component of various aspects of motivated behavior. Although mitogen-activated protein kinase (MAPK) signaling plays a crucial role in several types of learning, the cell type-specific role of MAPK pathway in stimulus-reward learning and motivation remains unclear. We herein investigated the role of MAPK in accumbal MSNs in reward-associated learning and memory. During the acquisition of Pavlovian conditioning, the number of phosphorylated MAPK1/3-positive cells was increased significantly and exclusively in the NAc core by 7-days of extensive training. MAPK signaling in the respective D1R- and D2R-MSNs was manipulated by transfecting an adeno-associated virus (AAV) plasmid into the NAc of Drd1a-Cre and Drd2-Cre transgenic mice. Potentiation of MAPK signaling shifted the learning curve of Pavlovian conditioning to the left only in Drd1a-Cre mice, whereas such manipulation in D2R-MSNs had negligible effects. In contrast, MAPK manipulation in D2R-MSNs of the NAc core significantly increased motivation for food rewards as found in Drd1a-Cre mice. These results suggest that MAPK signaling in the D1R-MSNs of NAc core plays an important role in stimulus-reward learning, while MAPK signaling in both D1R- and D2R-MSNs is involved in motivation for natural rewards.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
