

Signal transduction in astrocytes: Localization and release of inorganic polyphosphate
- PMID: 30260496
- PMCID: PMC6282517
- DOI: 10.1002/glia.23466
Signal transduction in astrocytes: Localization and release of inorganic polyphosphate
Abstract
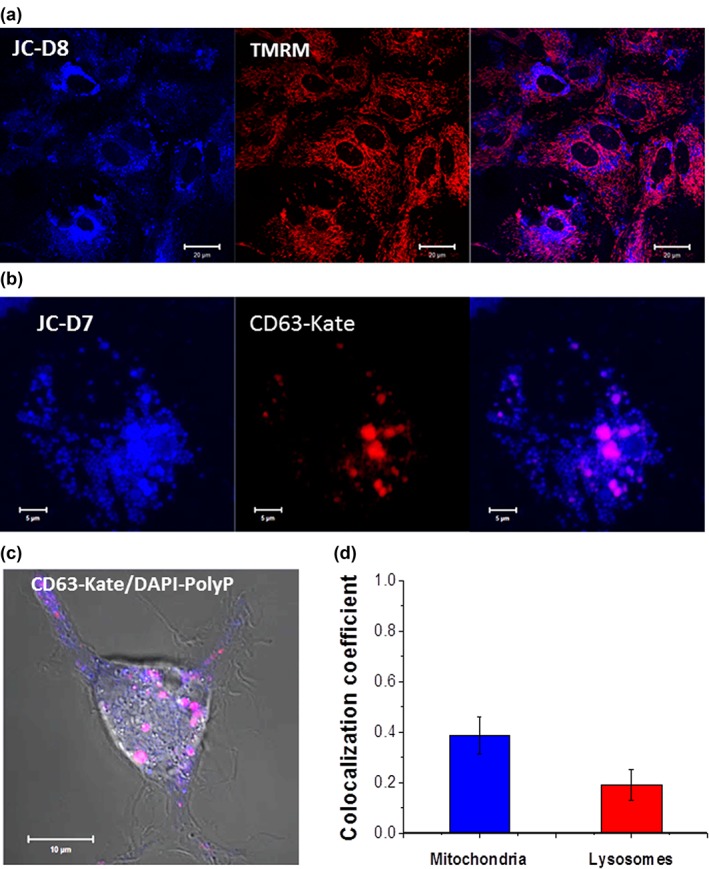
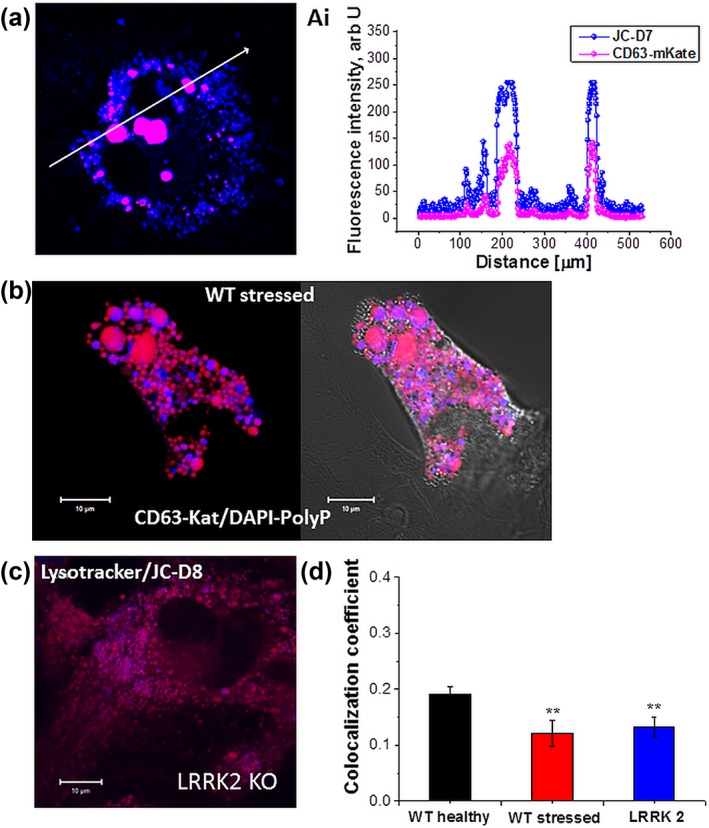
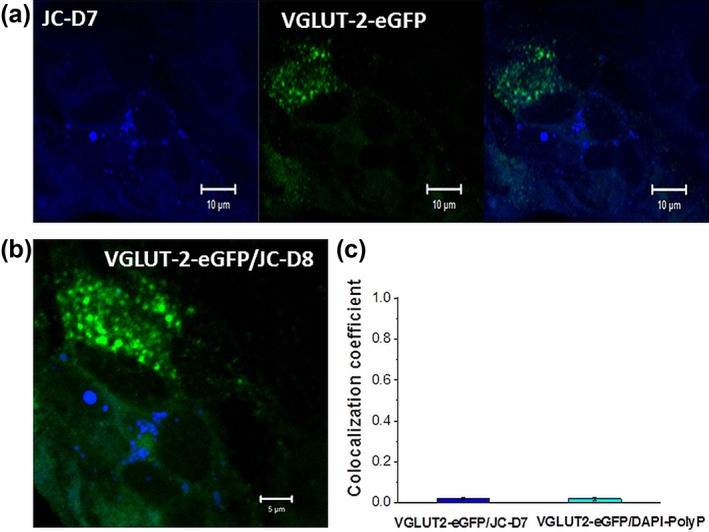
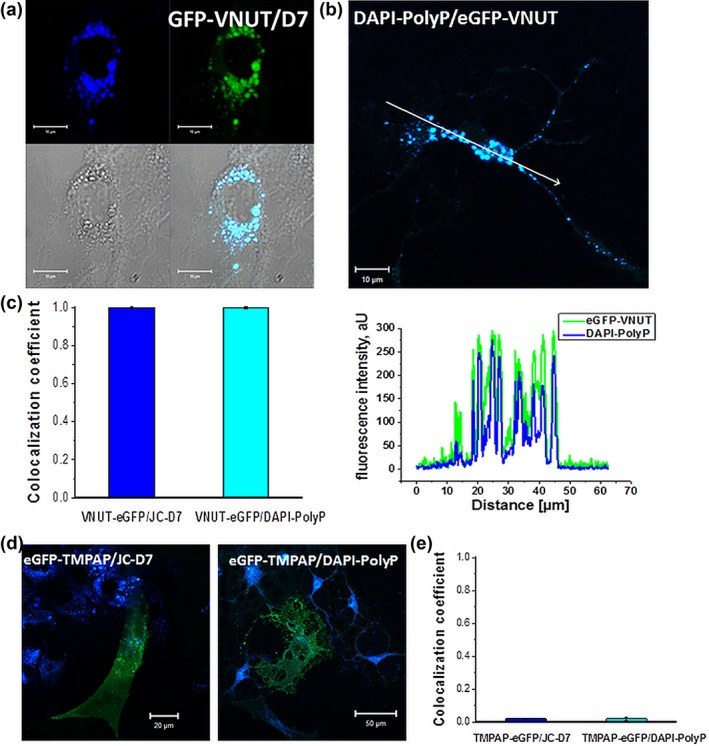
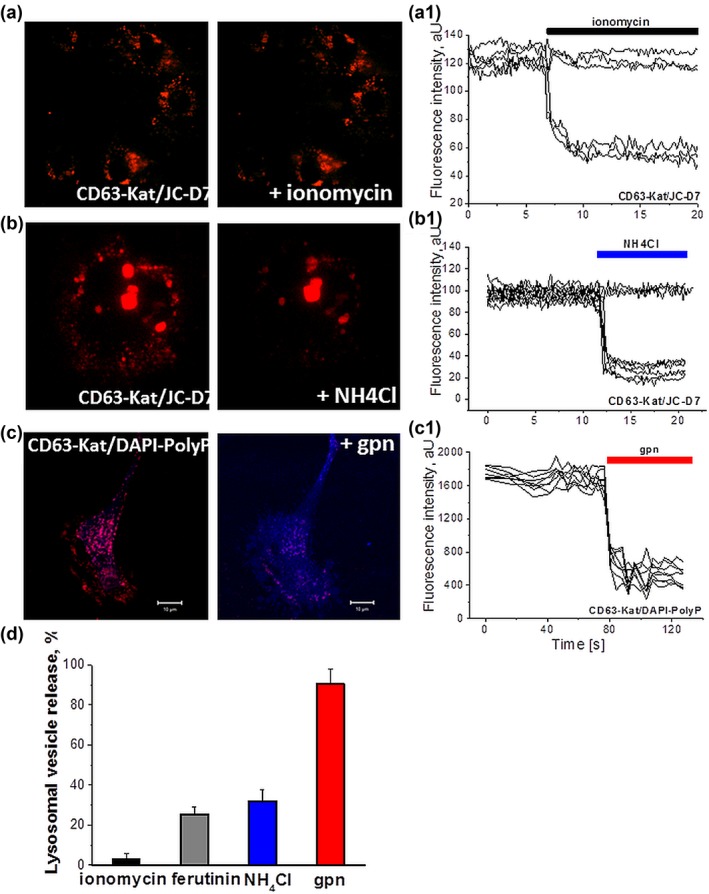
Inorganic polyphosphate (polyP) is present in every cell and is highly conserved from primeval times. In the mammalian cells, polyP plays multiple roles including control of cell bioenergetics and signal transduction. In the brain, polyP mediates signaling between astrocytes via activation of purinergic receptors, however, the mechanisms of polyP release remain unknown. Here we report identification of polyP-containing vesicles in cortical astrocytes and the main triggers that evoke vesicular polyP release. In cultured astrocytes, polyP was localized predominantly within the intracellular vesicular compartments which express vesicular nucleotide transporter VNUT (putative ATP-containing vesicles), but not within the compartments expressing vesicular glutamate transporter 2 (VGLUT2). The number of lysosomes which contain polyP was dependent on the conditions of astrocytes. Release of polyP from a proportion of lysosomes could be induced by calcium ionophores. In contrast, polyP release from the VNUT-containing vesicles could be triggered by various physiological stimuli, such as pH changes, polyP induced polyP release and other stimuli which increase [Ca2+ ] i . These data suggest that astrocytes release polyP predominantly via exocytosis from the VNUT-containing vesicles. © 2018 Wiley Periodicals, Inc.
Keywords: VNUT; astrocytes; inorganic polyphosphate; lysosomes; mitochondria.
© 2018 The Authors. Glia published by Wiley Periodicals, Inc.
Figures

Similar articles
-
Inorganic Polyphosphate Regulates AMPA and NMDA Receptors and Protects Against Glutamate Excitotoxicity via Activation of P2Y Receptors.J Neurosci. 2019 Jul 31;39(31):6038-6048. doi: 10.1523/JNEUROSCI.0314-19.2019. Epub 2019 May 30. J Neurosci. 2019. PMID: 31147524 Free PMC article.
-
Vesicular nucleotide transporter is involved in ATP storage of secretory lysosomes in astrocytes.Biochem Biophys Res Commun. 2013 Aug 16;438(1):145-51. doi: 10.1016/j.bbrc.2013.07.043. Epub 2013 Jul 19. Biochem Biophys Res Commun. 2013. PMID: 23876310
-
Signalling properties of inorganic polyphosphate in the mammalian brain.Nat Commun. 2013;4:1362. doi: 10.1038/ncomms2364. Nat Commun. 2013. PMID: 23322050 Free PMC article.
-
Vesicular nucleotide transporter (VNUT): appearance of an actress on the stage of purinergic signaling.Purinergic Signal. 2017 Sep;13(3):387-404. doi: 10.1007/s11302-017-9568-1. Epub 2017 Jun 14. Purinergic Signal. 2017. PMID: 28616712 Free PMC article. Review.
-
Role of inorganic polyphosphate in mammalian cells: from signal transduction and mitochondrial metabolism to cell death.Biochem Soc Trans. 2016 Feb;44(1):40-5. doi: 10.1042/BST20150223. Biochem Soc Trans. 2016. PMID: 26862186 Review.
Cited by
-
Inorganic Polyphosphate Regulates AMPA and NMDA Receptors and Protects Against Glutamate Excitotoxicity via Activation of P2Y Receptors.J Neurosci. 2019 Jul 31;39(31):6038-6048. doi: 10.1523/JNEUROSCI.0314-19.2019. Epub 2019 May 30. J Neurosci. 2019. PMID: 31147524 Free PMC article.
-
A Stringent Analysis of Polyphosphate Dynamics in Escherichia coli.J Bacteriol. 2019 Apr 9;201(9):e00070-19. doi: 10.1128/JB.00070-19. Print 2019 May 1. J Bacteriol. 2019. PMID: 30782636 Free PMC article.
-
The Reduced Level of Inorganic Polyphosphate Mobilizes Antioxidant and Manganese-Resistance Systems in Saccharomyces cerevisiae.Cells. 2019 May 15;8(5):461. doi: 10.3390/cells8050461. Cells. 2019. PMID: 31096715 Free PMC article.
-
Deletion of murine astrocytic vesicular nucleotide transporter increases anxiety and depressive-like behavior and attenuates motivation for reward.Mol Psychiatry. 2025 Feb;30(2):506-520. doi: 10.1038/s41380-024-02692-5. Epub 2024 Aug 9. Mol Psychiatry. 2025. PMID: 39122778 Free PMC article.
-
Astrocytes in cocaine addiction and beyond.Mol Psychiatry. 2022 Jan;27(1):652-668. doi: 10.1038/s41380-021-01080-7. Epub 2021 Apr 9. Mol Psychiatry. 2022. PMID: 33837268 Free PMC article. Review.
References
-
- Abramov, A. Y. , & Duchen, M. R. (2003). Actions of ionomycin, 4‐BrA23187 and a novel electrogenic Ca2+ ionophore on mitochondria in intact cells. Cell Calcium, 33(2), 101–112. S0143416002002038 [pii] - PubMed
-
- Abramov, A. Y. , Fraley, C. , Diao, C. T. , Winkfein, R. , Colicos, M. A. , Duchen, M. R. , … Pavlov, E. (2007). Targeted polyphosphatase expression alters mitochondrial metabolism and inhibits calcium‐dependent cell death. Proceedings of the National Academy of Sciences of the United States of America, 104(46), 18091–18096. doi:10.1073/pnas.0708959104 [doi] - PMC - PubMed
-
- Abramov, A. Y. , Zamaraeva, M. V. , Hagelgans, A. I. , Azimov, R. R. , & Krasilnikov, O. V. (2001). Influence of plant terpenoids on the permeability of mitochondria and lipid bilayers. Biochimica et Biophysica Acta, 1512(1), 98–110. S0005‐2736(01)00307‐8 [pii] - PubMed
-
- Angelova, P. R. , & Abramov, A. Y. (2014). Interaction of neurons and astrocytes underlies the mechanism of Abeta‐induced neurotoxicity. Biochemical Society Transactions, 42(5), 1286–1290. doi:10.1042/BST20140153 - PubMed
-
- Angelova, P. R. , Agrawalla, B. K. , Elustondo, P. A. , Gordon, J. , Shiba, T. , Abramov, A. Y. , … Pavlov, E. V. (2014). In situ investigation of mammalian inorganic polyphosphate localization using novel selective fluorescent probes JC‐D7 and JC‐D8. ACS Chemical Biology, 9(9), 2101–2110. doi:10.1021/cb5000696 [doi] - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
