

Mapping the Contact Sites of the Escherichia coli Division-Initiating Proteins FtsZ and ZapA by BAMG Cross-Linking and Site-Directed Mutagenesis
- PMID: 30261644
- PMCID: PMC6213154
- DOI: 10.3390/ijms19102928
Mapping the Contact Sites of the Escherichia coli Division-Initiating Proteins FtsZ and ZapA by BAMG Cross-Linking and Site-Directed Mutagenesis
Abstract
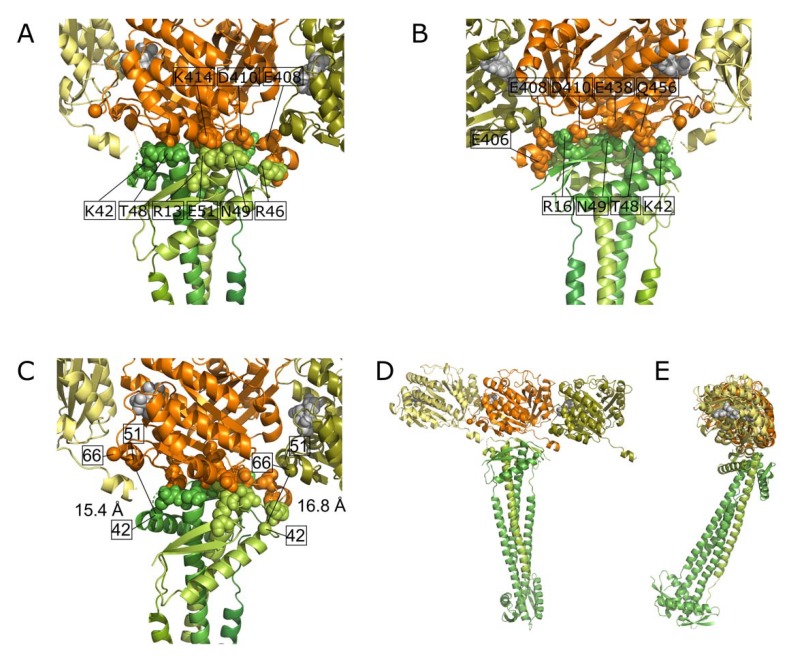
Cell division in bacteria is initiated by the polymerization of FtsZ at midcell in a ring-like structure called the Z-ring. ZapA and other proteins assist Z-ring formation and ZapA binds ZapB, which senses the presence of the nucleoids. The FtsZ⁻ZapA binding interface was analyzed by chemical cross-linking mass spectrometry (CXMS) under in vitro FtsZ-polymerizing conditions in the presence of GTP. Amino acids residue K42 from ZapA was cross-linked to amino acid residues K51 and K66 from FtsZ, close to the interphase between FtsZ molecules in protofilaments. Five different cross-links confirmed the tetrameric structure of ZapA. A number of FtsZ cross-links suggests that its C-terminal domain of 55 residues, thought to be largely disordered, has a limited freedom to move in space. Site-directed mutagenesis of ZapA reveals an interaction site in the globular head of the protein close to K42. Using the information on the cross-links and the mutants that lost the ability to interact with FtsZ, a model of the FtsZ protofilament⁻ZapA tetramer complex was obtained by information-driven docking with the HADDOCK2.2 webserver.
Keywords: 1,4-bis(succimidyl)-3-azidomethylglutarate (BAMG); Filamenting temperature sensitive Z (FtsZ); Fourier-Transform Ion Cyclotron Resonance mass spectrometry(FTICR); Z associated protein A (ZapA); cell division; quadrupole time of flight mass spectrometer (QTOF).
Conflict of interest statement
The authors declare no conflict of interest.
Figures
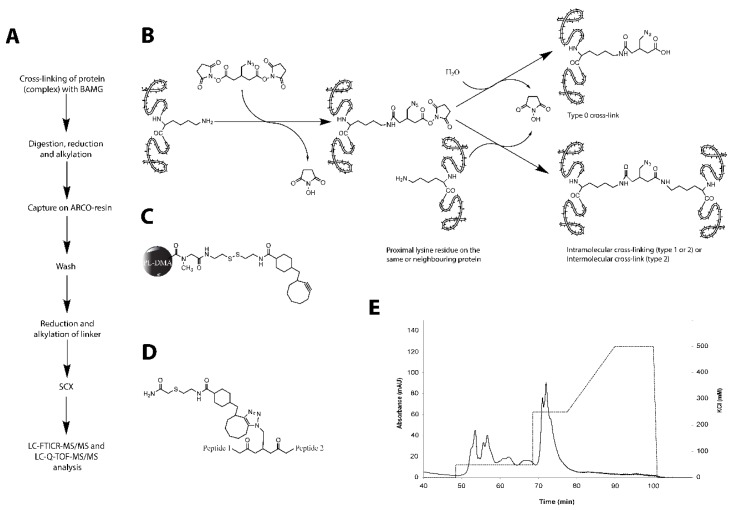
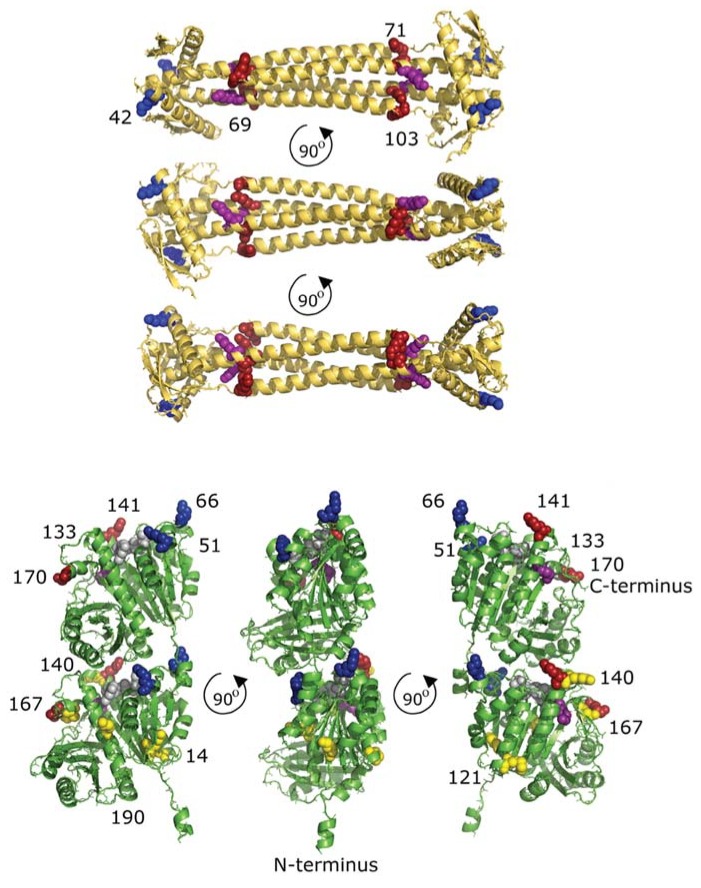
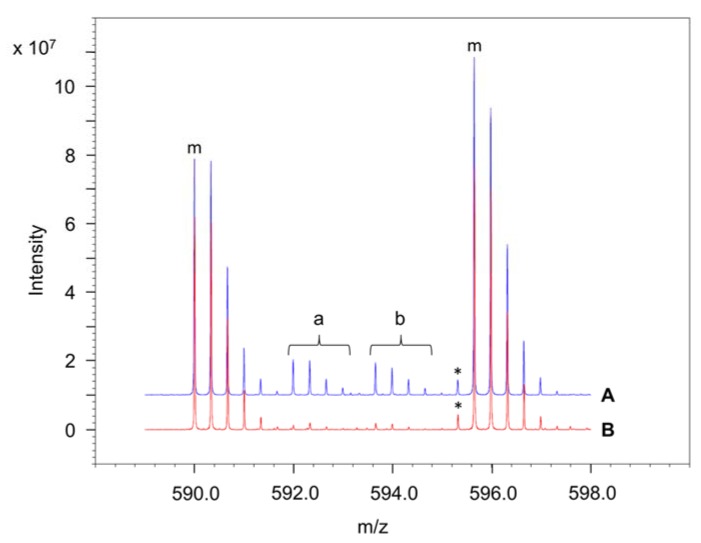
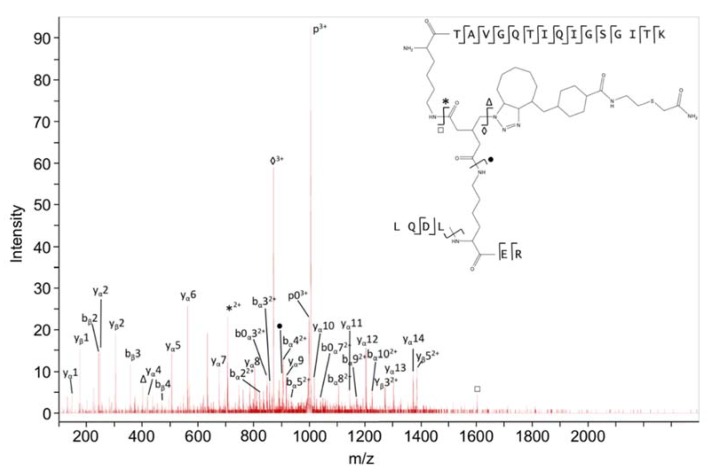

Similar articles
-
Structure and Mutational Analyses of Escherichia coli ZapD Reveal Charged Residues Involved in FtsZ Filament Bundling.J Bacteriol. 2016 May 13;198(11):1683-1693. doi: 10.1128/JB.00969-15. Print 2016 Jun 1. J Bacteriol. 2016. PMID: 27021560 Free PMC article.
-
ZapA and ZapB form an FtsZ-independent structure at midcell.Mol Microbiol. 2017 May;104(4):652-663. doi: 10.1111/mmi.13655. Epub 2017 Mar 26. Mol Microbiol. 2017. PMID: 28249098 Free PMC article.
-
Cross-linking FtsZ polymers into coherent Z rings.Mol Microbiol. 2010 Nov;78(3):651-68. doi: 10.1111/j.1365-2958.2010.07352.x. Epub 2010 Sep 20. Mol Microbiol. 2010. PMID: 20969647
-
Strong FtsZ is with the force: mechanisms to constrict bacteria.Trends Microbiol. 2010 Aug;18(8):348-56. doi: 10.1016/j.tim.2010.06.001. Epub 2010 Jul 1. Trends Microbiol. 2010. PMID: 20598544 Review.
-
The keepers of the ring: regulators of FtsZ assembly.FEMS Microbiol Rev. 2016 Jan;40(1):57-67. doi: 10.1093/femsre/fuv040. Epub 2015 Sep 15. FEMS Microbiol Rev. 2016. PMID: 26377318 Review.
Cited by
-
Regulation of cytokinesis: FtsZ and its accessory proteins.Curr Genet. 2020 Feb;66(1):43-49. doi: 10.1007/s00294-019-01005-6. Epub 2019 Jun 17. Curr Genet. 2020. PMID: 31209564 Review.
-
Indole-core-based novel antibacterial agent targeting FtsZ.Infect Drug Resist. 2019 Jul 24;12:2283-2296. doi: 10.2147/IDR.S208757. eCollection 2019. Infect Drug Resist. 2019. PMID: 31413605 Free PMC article.
-
ZapG (YhcB/DUF1043), a novel cell division protein in gamma-proteobacteria linking the Z-ring to septal peptidoglycan synthesis.J Biol Chem. 2021 Jan-Jun;296:100700. doi: 10.1016/j.jbc.2021.100700. Epub 2021 Apr 23. J Biol Chem. 2021. PMID: 33895137 Free PMC article.
-
A Novel Z-Ring Associated Protein ZapA-Like Protein (PA5407) From Pseudomonas aeruginosa Promotes FtsZ to Form Double Filaments.Front Microbiol. 2021 Aug 4;12:717013. doi: 10.3389/fmicb.2021.717013. eCollection 2021. Front Microbiol. 2021. PMID: 34421877 Free PMC article.
-
Transient Membrane-Linked FtsZ Assemblies Precede Z-Ring Formation in Escherichia coli.Curr Biol. 2020 Feb 3;30(3):499-508.e6. doi: 10.1016/j.cub.2019.12.023. Epub 2020 Jan 23. Curr Biol. 2020. PMID: 31978334 Free PMC article.
References
-
- Van der Ploeg R., Verheul J., Vischer N.O.E., Alexeeva S., Hoogendoorn E., Postma M., Banzhaf M., Vollmer W., den Blaauwen T. Colocalization and interaction between elongasome and divisome during a preparative cell division phase in Escherichia coli. Mol. Microbiol. 2013;87:1074–1087. doi: 10.1111/mmi.12150. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
