

Single-cell analysis uncovers convergence of cell identities during axolotl limb regeneration
- PMID: 30262634
- PMCID: PMC6669047
- DOI: 10.1126/science.aaq0681
Single-cell analysis uncovers convergence of cell identities during axolotl limb regeneration
Abstract
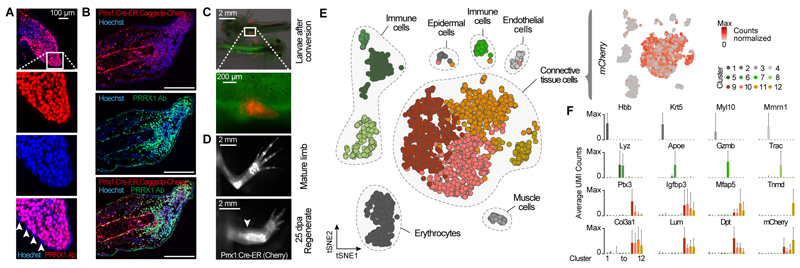
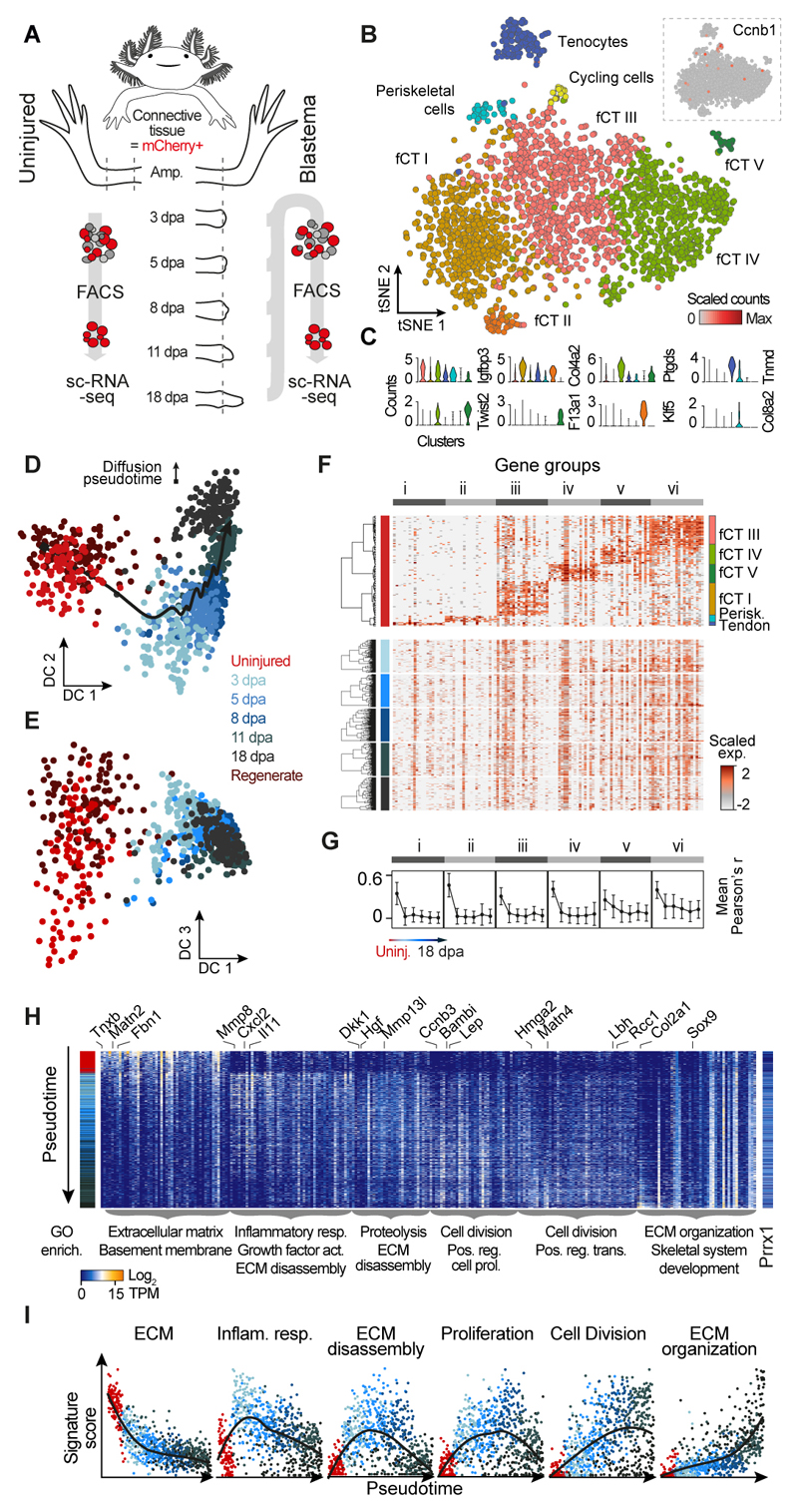
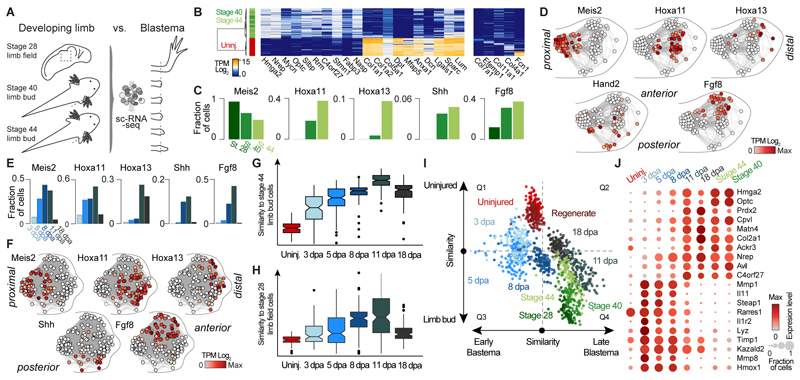
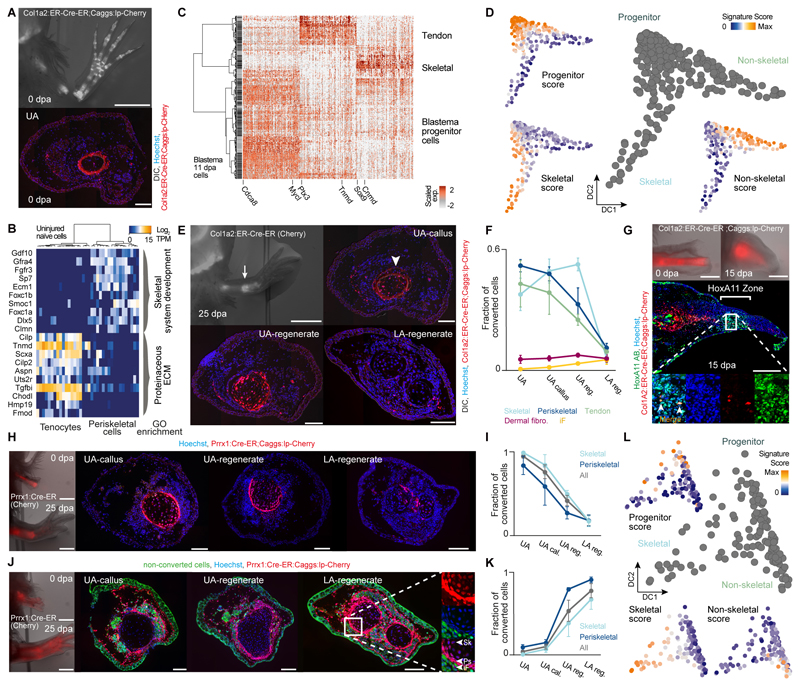
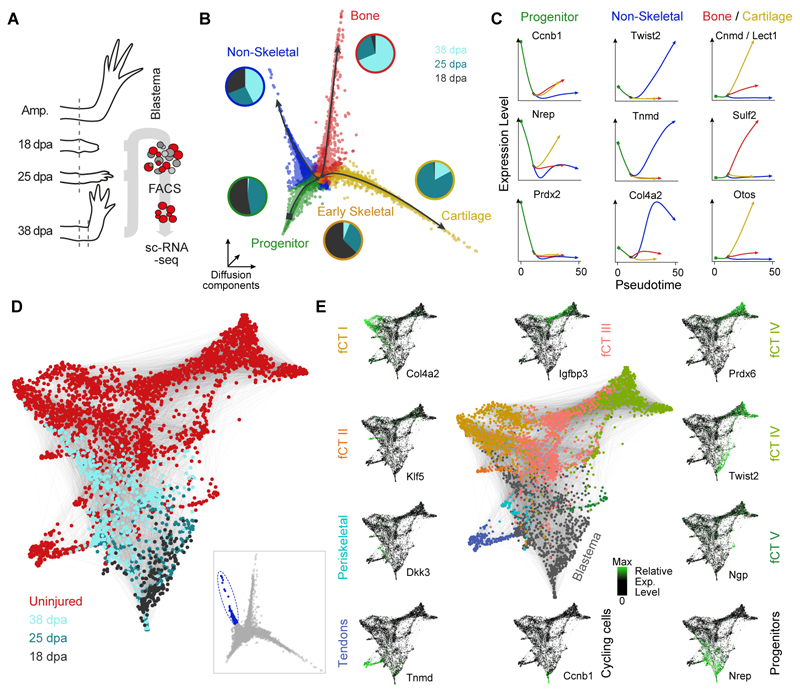
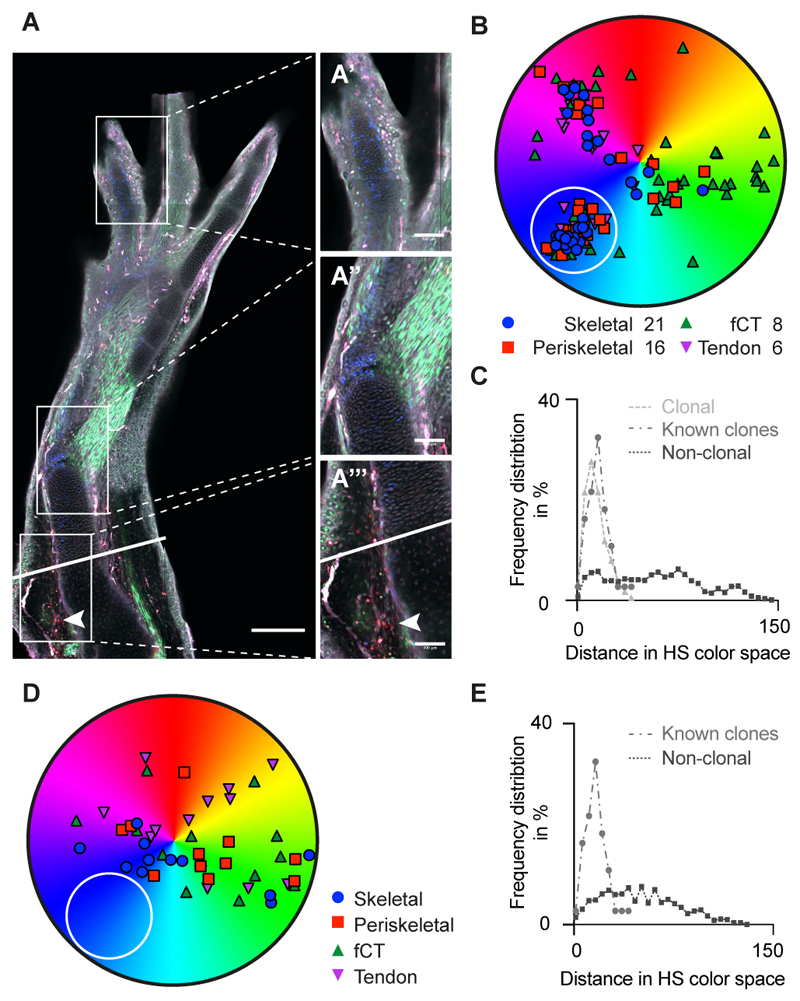
Amputation of the axolotl forelimb results in the formation of a blastema, a transient tissue where progenitor cells accumulate prior to limb regeneration. However, the molecular understanding of blastema formation had previously been hampered by the inability to identify and isolate blastema precursor cells in the adult tissue. We have used a combination of Cre-loxP reporter lineage tracking and single-cell messenger RNA sequencing (scRNA-seq) to molecularly track mature connective tissue (CT) cell heterogeneity and its transition to a limb blastema state. We have uncovered a multiphasic molecular program where CT cell types found in the uninjured adult limb revert to a relatively homogenous progenitor state that recapitulates an embryonic limb bud-like phenotype including multipotency within the CT lineage. Together, our data illuminate molecular and cellular reprogramming during complex organ regeneration in a vertebrate.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
Comment in
-
Tracing the Origins of Axolotl Limb Regeneration.Dev Cell. 2018 Dec 17;47(6):675-677. doi: 10.1016/j.devcel.2018.11.042. Dev Cell. 2018. PMID: 30562504
References
-
- Brockes JP, Kumar A. Comparative aspects of animal regeneration. Annu Rev Cell Dev Biol. 2008;24:525–549. - PubMed
-
- Nacu E, Tanaka EM. Limb regeneration: a new development? Annu Rev Cell Dev Biol. 2011;27:409–440. - PubMed
-
- Muneoka K, Fox WF, Bryant SV. Cellular contribution from dermis and cartilage to the regenerating limb blastema in axolotls. Dev Biol. 1986;116:256–260. - PubMed
-
- Kragl M, et al. Cells keep a memory of their tissue origin during axolotl limb regeneration. Nature. 2009;460:60–65. - PubMed
-
- Dunis DA, Namenwirth M. The role of grafted skin in the regeneration of x-irradiated axolotl limbs. Dev Biol. 1977;56:97–109. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
