

Hallmarks of primate lentiviral immunodeficiency infection recapitulate loss of innate lymphoid cells
- PMID: 30262807
- PMCID: PMC6160474
- DOI: 10.1038/s41467-018-05528-3
Hallmarks of primate lentiviral immunodeficiency infection recapitulate loss of innate lymphoid cells
Abstract
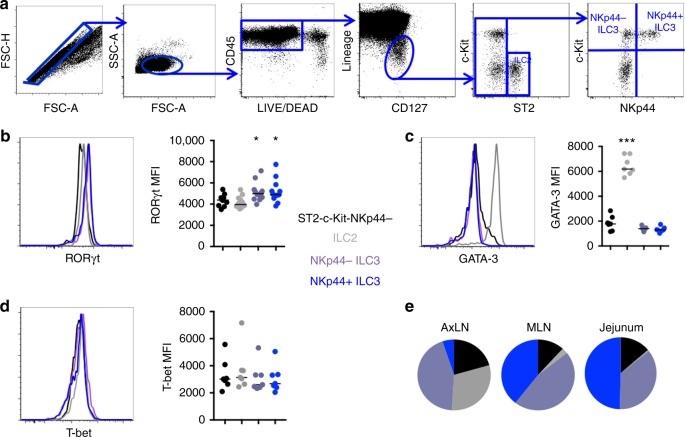
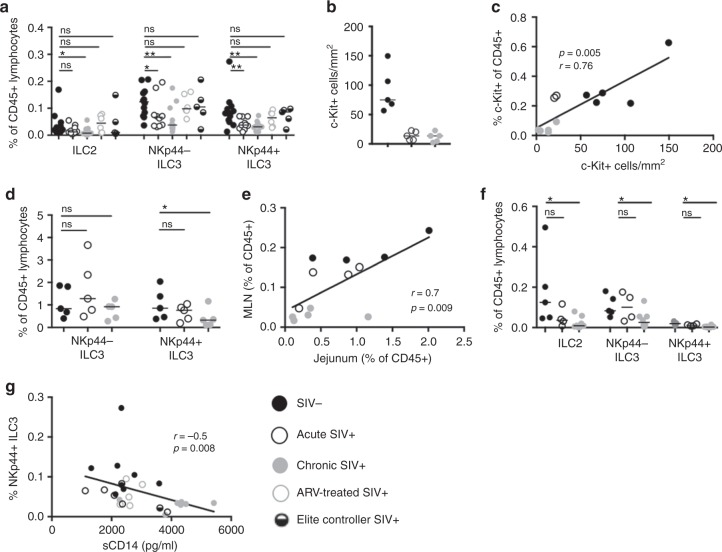
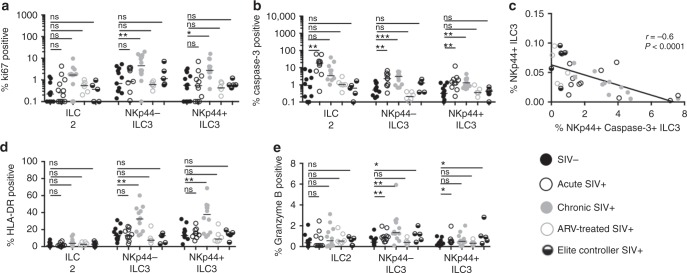
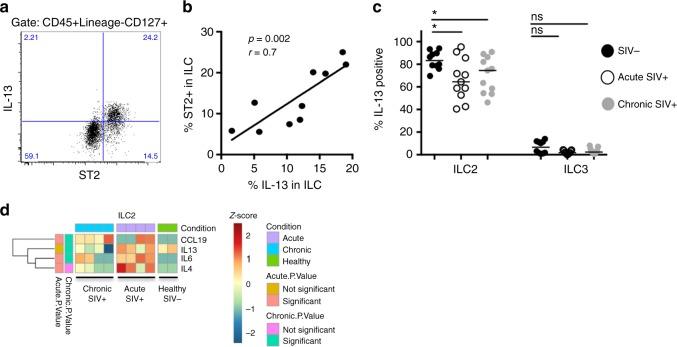
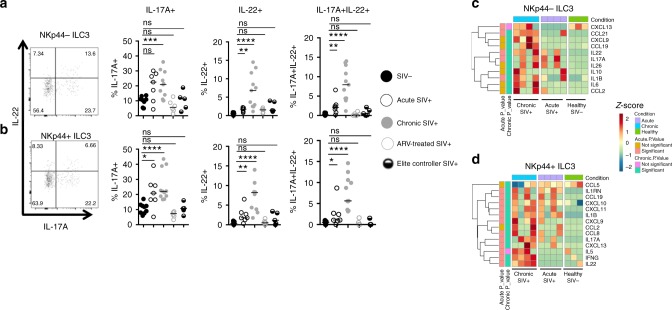
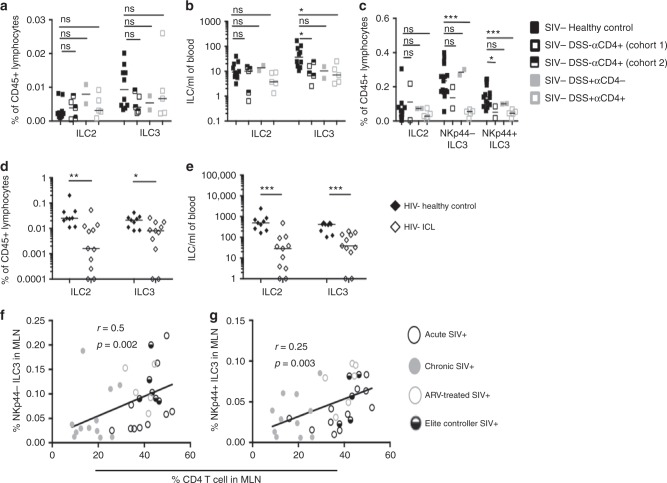
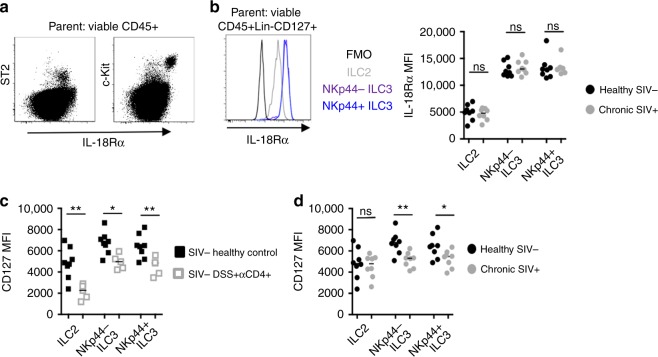
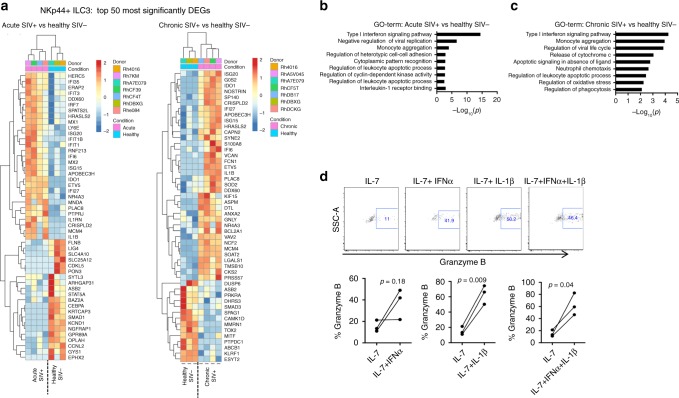
Innate lymphoid cells (ILCs) play critical roles in mucosal barrier defense and tissue homeostasis. While ILCs are depleted in HIV-1 infection, this phenomenon is not a generalized feature of all viral infections. Here we show in untreated SIV-infected rhesus macaques (RMs) that ILC3s are lost rapidly in mesenteric lymph nodes (MLNs), yet preserved in SIV+ RMs with pharmacologic or natural control of viremia. In healthy uninfected RMs, experimental depletion of CD4+ T cells in combination with dextran sodium sulfate (DSS) is sufficient to reduce ILC frequencies in the MLN. In this setting and in chronic SIV+ RMs, IL-7Rα chain expression diminishes on ILC3s in contrast to the IL-18Rα chain expression which remains stable. In HIV-uninfected patients with durable CD4+ T cell deficiency (deemed idiopathic CD4+ lymphopenia), similar ILC deficiencies in blood were observed, collectively identifying determinants of ILC homeostasis in primates and potential mechanisms underlying their depletion in HIV/SIV infection.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Presence of Inflammatory Group I and III Innate Lymphoid Cells in the Colon of Simian Immunodeficiency Virus-Infected Rhesus Macaques.J Virol. 2020 Apr 16;94(9):e01914-19. doi: 10.1128/JVI.01914-19. Print 2020 Apr 16. J Virol. 2020. PMID: 32051277 Free PMC article.
-
Functional Perturbation of Mucosal Group 3 Innate Lymphoid and Natural Killer Cells in Simian-Human Immunodeficiency Virus/Simian Immunodeficiency Virus-Infected Infant Rhesus Macaques.J Virol. 2020 Feb 14;94(5):e01644-19. doi: 10.1128/JVI.01644-19. Print 2020 Feb 14. J Virol. 2020. PMID: 31801861 Free PMC article.
-
Differential Dynamics of Regulatory T-Cell and Th17 Cell Balance in Mesenteric Lymph Nodes and Blood following Early Antiretroviral Initiation during Acute Simian Immunodeficiency Virus Infection.J Virol. 2019 Sep 12;93(19):e00371-19. doi: 10.1128/JVI.00371-19. Print 2019 Oct 1. J Virol. 2019. PMID: 31315987 Free PMC article.
-
Innate Lymphoid Cells: Their Contributions to Gastrointestinal Tissue Homeostasis and HIV/SIV Disease Pathology.Curr HIV/AIDS Rep. 2019 Jun;16(3):181-190. doi: 10.1007/s11904-019-00439-4. Curr HIV/AIDS Rep. 2019. PMID: 31104270 Free PMC article. Review.
-
The Hitchhiker Guide to CD4+ T-Cell Depletion in Lentiviral Infection. A Critical Review of the Dynamics of the CD4+ T Cells in SIV and HIV Infection.Front Immunol. 2021 Jul 21;12:695674. doi: 10.3389/fimmu.2021.695674. eCollection 2021. Front Immunol. 2021. PMID: 34367156 Free PMC article.
Cited by
-
SIV infection and ARV treatment reshape the transcriptional and epigenetic profile of naïve and memory T cells in vivo.J Virol. 2024 Jun 13;98(6):e0028324. doi: 10.1128/jvi.00283-24. Epub 2024 May 23. J Virol. 2024. PMID: 38780248 Free PMC article.
-
Functional interactions between innate lymphoid cells and adaptive immunity.Nat Rev Immunol. 2019 Oct;19(10):599-613. doi: 10.1038/s41577-019-0194-8. Epub 2019 Jul 26. Nat Rev Immunol. 2019. PMID: 31350531 Free PMC article. Review.
-
Combined protein and nucleic acid imaging reveals virus-dependent B cell and macrophage immunosuppression of tissue microenvironments.Immunity. 2022 Jun 14;55(6):1118-1134.e8. doi: 10.1016/j.immuni.2022.03.020. Epub 2022 Apr 20. Immunity. 2022. PMID: 35447093 Free PMC article.
-
Biomarkers of Cellular Stress Do Not Associate with sCD14 in Progressive HIV and SIV Infections in Vivo.Pathog Immun. 2020 Apr 24;5(1):68-88. doi: 10.20411/pai.v5i1.363. eCollection 2020. Pathog Immun. 2020. PMID: 32426577 Free PMC article.
-
Translocating bacteria in SIV infection are not stochastic and preferentially express cytosine methyltransferases.Mucosal Immunol. 2024 Oct;17(5):1089-1101. doi: 10.1016/j.mucimm.2024.07.008. Epub 2024 Jul 31. Mucosal Immunol. 2024. PMID: 39089468 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
