

GNA13 promotes tumor growth and angiogenesis by upregulating CXC chemokines via the NF-κB signaling pathway in colorectal cancer cells
- PMID: 30267476
- PMCID: PMC6246959
- DOI: 10.1002/cam4.1783
GNA13 promotes tumor growth and angiogenesis by upregulating CXC chemokines via the NF-κB signaling pathway in colorectal cancer cells
Retraction in
-
Retraction.Cancer Med. 2023 Sep;12(18):19352. doi: 10.1002/cam4.6367. Epub 2023 Sep 19. Cancer Med. 2023. PMID: 37724781 Free PMC article.
Abstract
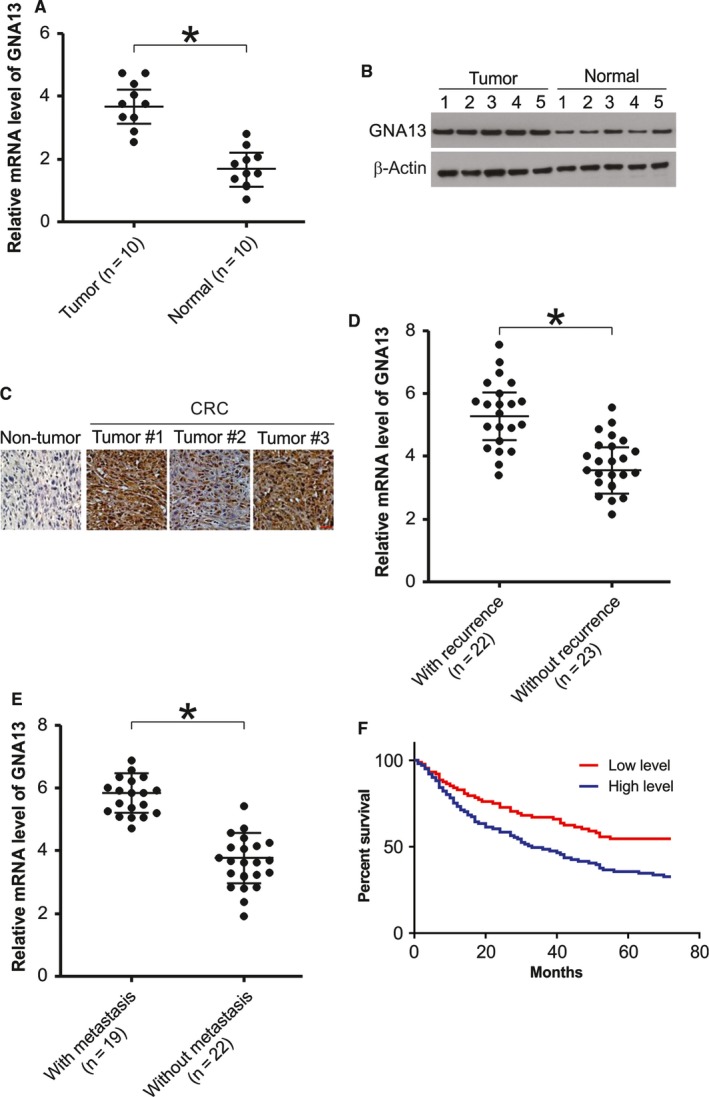
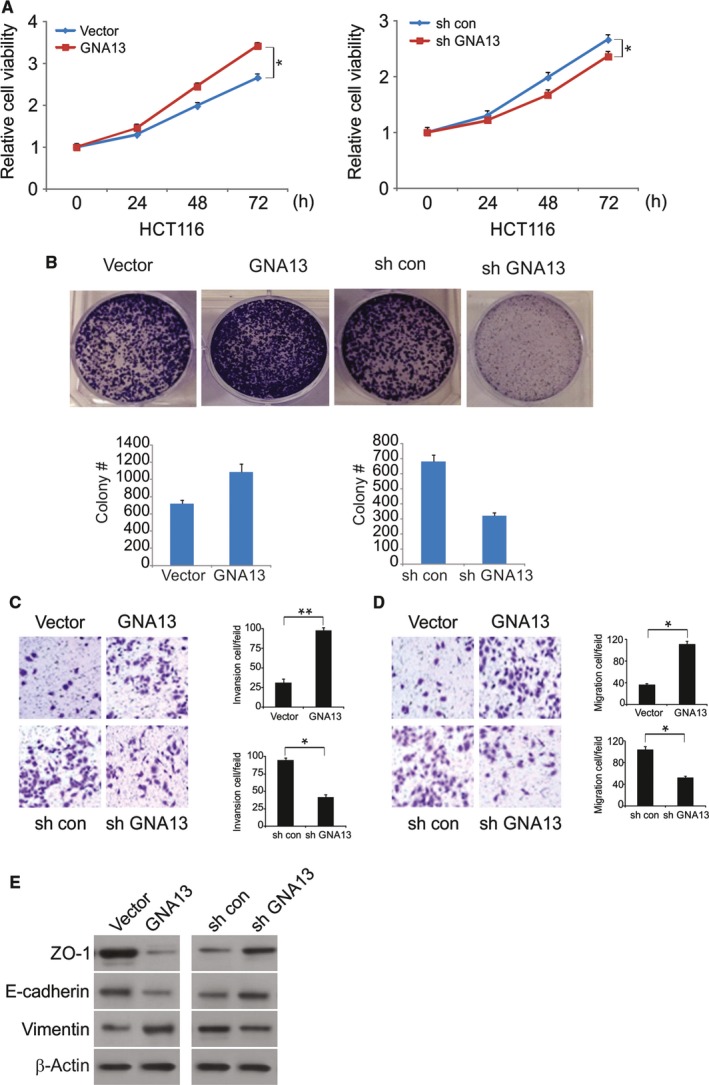
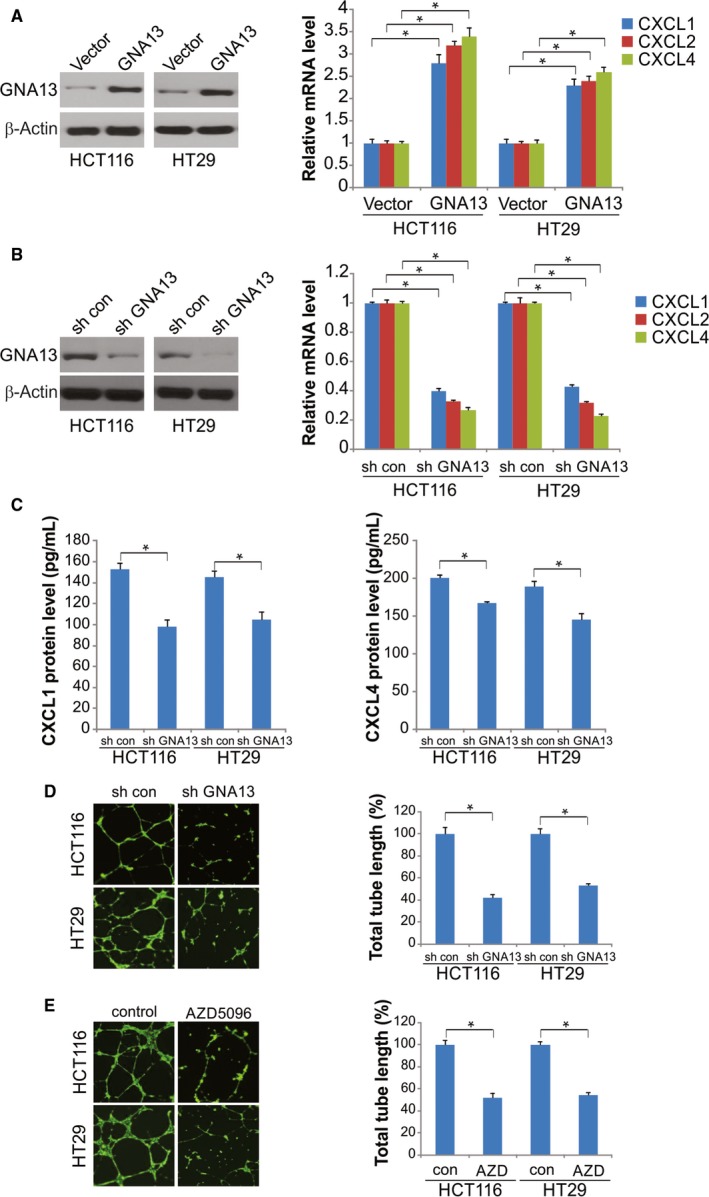
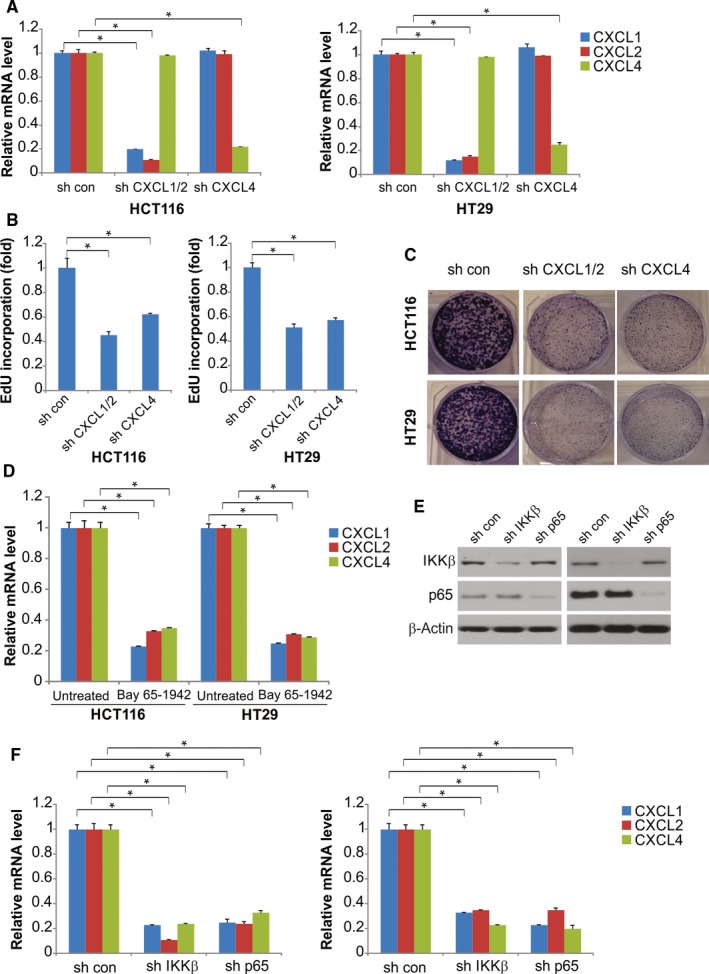
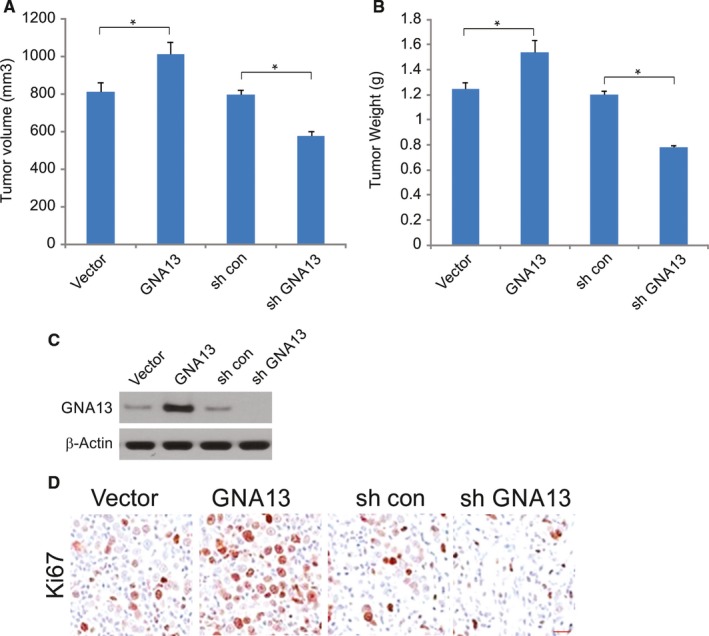
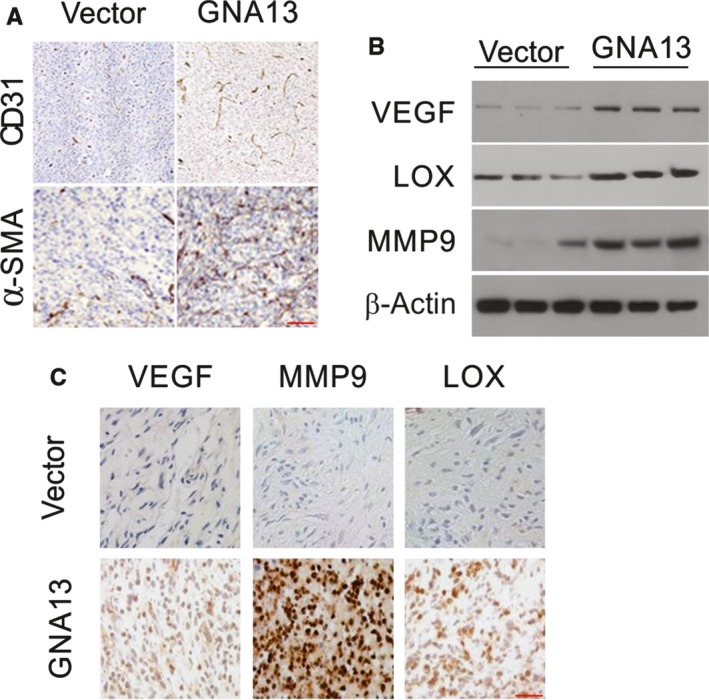
GNA13 has been found overexpressed in various types of cancer, which is related to tumor metastasis and progression. However, the biological functions of GNA13 in colorectal cancer (CRC) progression remain unclear. This study aimed to explore the role of GNA13 in CRC and investigate the mechanism of how GNA13 promotes tumor growth. Interestingly, our findings showed that GNA13 is commonly upregulated in CRC, where these events are associated with a worse histologic grade and poor survival. Increased expression levels of GNA13 promoted cell growth, migration, invasion, and epithelial-mesenchymal transition, whereas GNA13 silencing abrogated these malignant phenotypes. In addition, overexpressing GNA13 in cancer cells increased the levels of the chemokines CXCL1, CXCL2, and CXCL4, which contributed to CRC proliferation and colony formation. Moreover, our mechanistic investigations suggest that the NF-κB/p65 signaling pathway was activated by the increase in GNA13 levels. Inhibiting the NF-κB/p65 pathway with an inhibitor decreased GNA13-induced migration, invasion and CXCL chemokine level increases, indicating the critical role of NF-κB/p65 signaling in mediating the effects of GNA13 in CRC. Together, these results demonstrate a key role of GNA13 overexpression in CRC that contributes to malignant behavior in cancer cells, at least in part through stimulating angiogenesis and increasing the levels of the NF-κB-dependent chemokines CXCL1, CXCL2, and CXCL4.
Keywords: CRC; CXCL chemokines; GNA13; NF-κB.
© 2018 The Authors. Cancer Medicine published by John Wiley & Sons Ltd.
Figures
References
-
- Wu F, Song G, de Graaf C, Stevens RC. Structure and function of peptide‐binding G protein‐coupled receptors. J Mol Biol. 2017;429:2726‐2745. - PubMed
-
- Latek D, Bajda M, Filipek S. A hybrid approach to structure and function modeling of G protein‐coupled receptors. J Chem Inf Model. 2016;56:630‐641. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
