

Comparative Analysis of cul5 and rbx2 Expression in the Developing and Adult Murine Brain and Their Essentiality During Mouse Embryogenesis
- PMID: 30269386
- PMCID: PMC6639038
- DOI: 10.1002/dvdy.24675
Comparative Analysis of cul5 and rbx2 Expression in the Developing and Adult Murine Brain and Their Essentiality During Mouse Embryogenesis
Abstract
Background: The E3 Cullin 5-RING ubiquitin ligase (CRL5) is a multiprotein complex that has recently been highlighted as a major regulator of central nervous system development. Cullin 5 (Cul5) and the RING finger protein Rbx2 are two CRL5 core components required for CRL5 function in the brain, but their full expression patterns and developmental functions have not been described in detail.
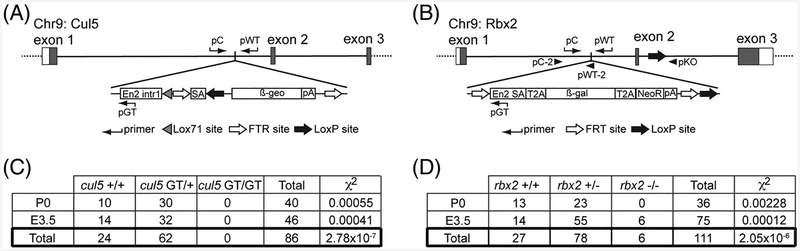
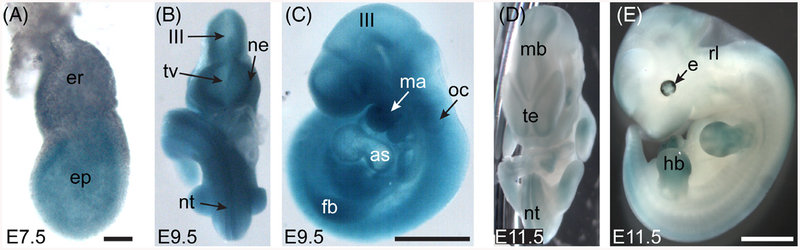
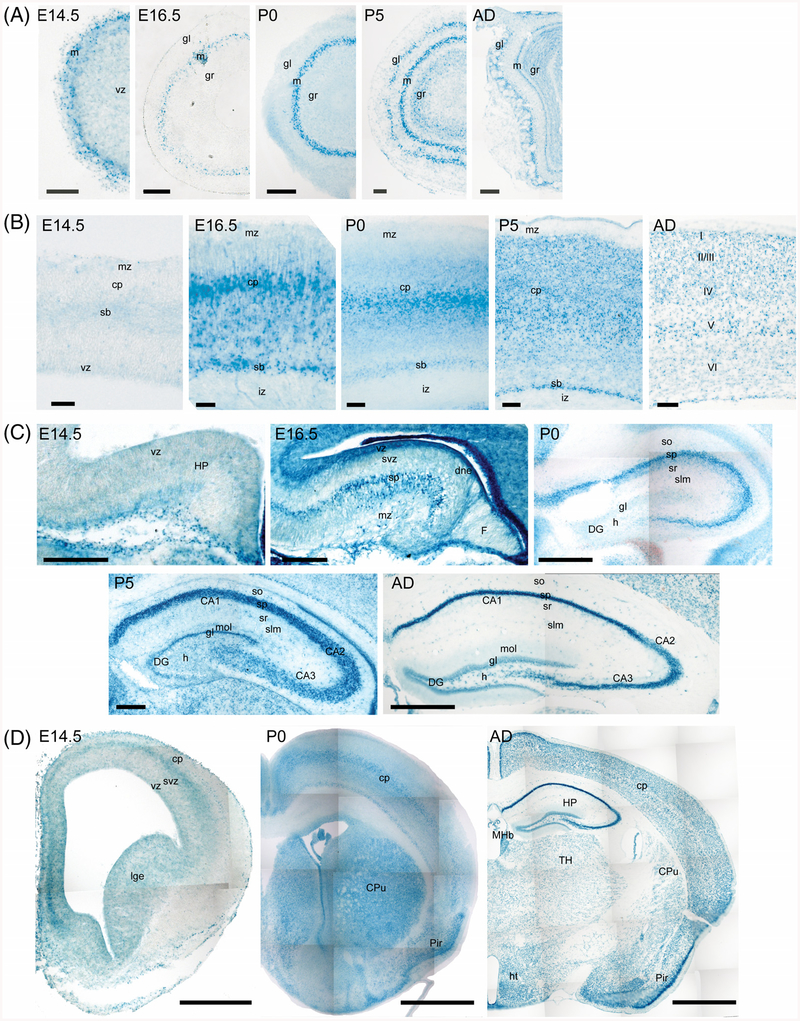
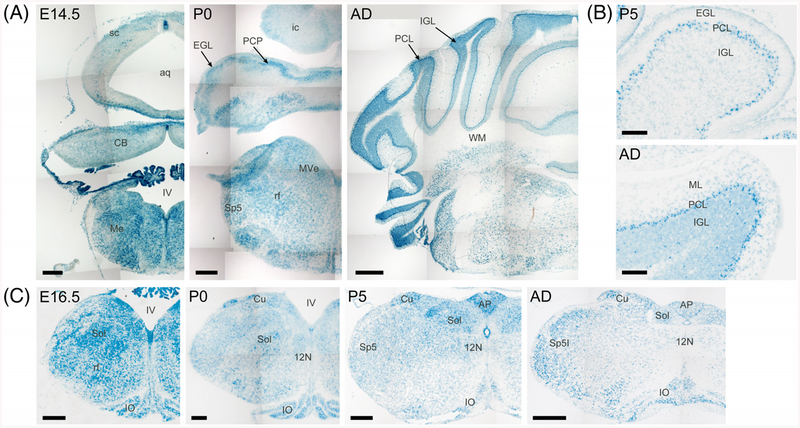
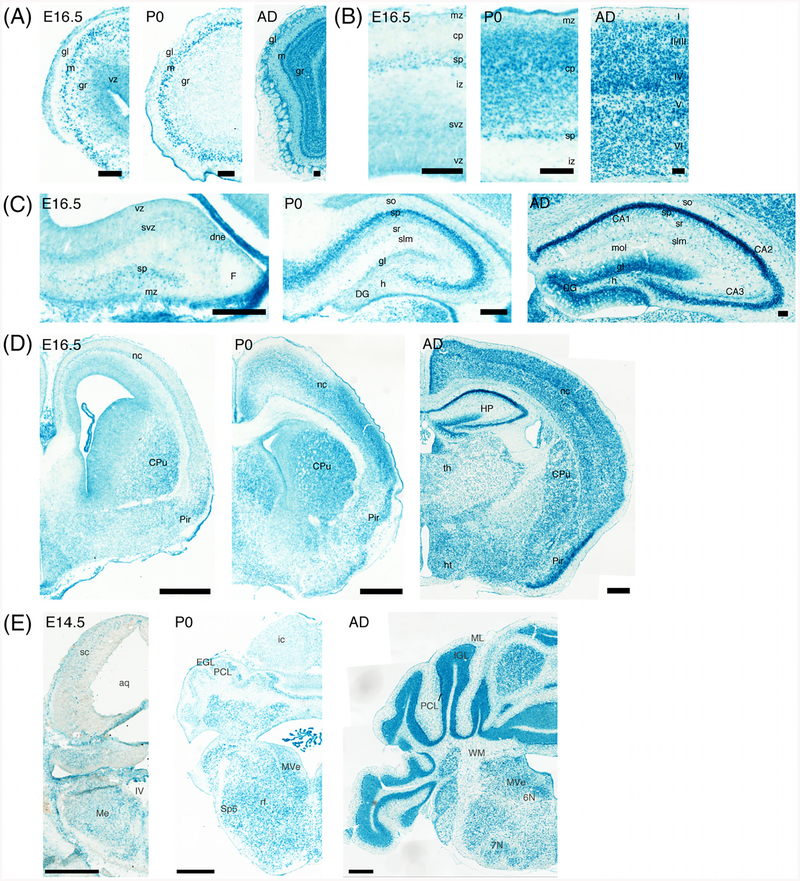
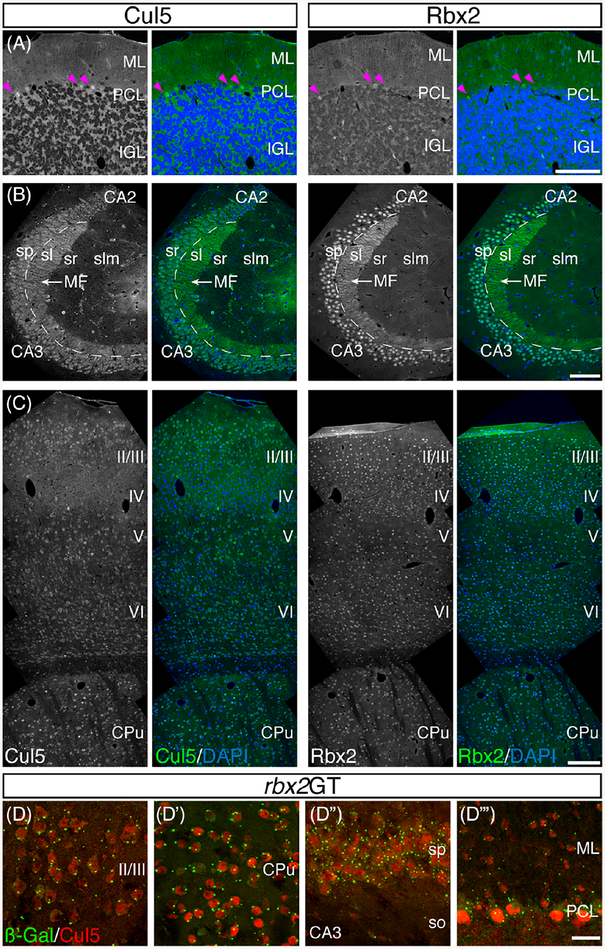
Results: Using a gene-trap mouse model for Cul5 and a knock-in-knockout mouse model for Rbx2, we show that lack of Cul5, but not Rbx2, disrupts blastocyst formation. However, Rbx2 is required for embryo survival at later embryonic stages. We also show that cul5 is expressed in the embryo proper as early as E7.5 and its expression is mostly restricted to the central nervous system and limbs at later time points. Finally, we show that rbx2 and cul5 are co-expressed in most areas of the brain during development and in the adult.
Conclusions: Our results show that Cul5, but not Rbx2, is required during early embryogenesis and suggests that Cul5 has Rbx2-independent functions in early development. In the brain, Cul5 and Rbx2 are expressed in a similar fashion, allowing the nucleation of an active CRL5 complex. Developmental Dynamics 247:1227-1236, 2018. © 2018 Wiley Periodicals, Inc.
Keywords: cul5; rbx2; CRL5; Embryogenesis; brain development.
© 2018 Wiley Periodicals, Inc.
Figures
References
-
- Bayer SA. 1983. 3H-thymidine-radiographic studies of neurogenesis in the rat olfactory bulb. Exp Brain Res 50:329–340. - PubMed
-
- Ceremuga TE, Yao XL, McCabe JT. 2001. Vasopressin-activated calcium-mobilizing (VACM-1) receptor mRNA is present in peripheral organs and the central nervous system of the laboratory rat. Endocr Res 27:433–445. - PubMed
-
- Ceremuga TE, Yao XL, McCabe JT. 2003. Cullin-5 is ubiquitous in the rat brain. Neurosci Lett 345:121–125. - PubMed
-
- Dealy MJ, Nguyen KV, Lo J, Gstaiger M, Krek W, Elson D, Arbeit J, Kipreos ET, Johnson RS. 1999. Loss of Cul1 results in early embryonic lethality and dysregulation of cyclin E. Nat Genet 23:245–248. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
