

Protein AMPylation by an Evolutionarily Conserved Pseudokinase
- PMID: 30270044
- PMCID: PMC6524645
- DOI: 10.1016/j.cell.2018.08.046
Protein AMPylation by an Evolutionarily Conserved Pseudokinase
Abstract
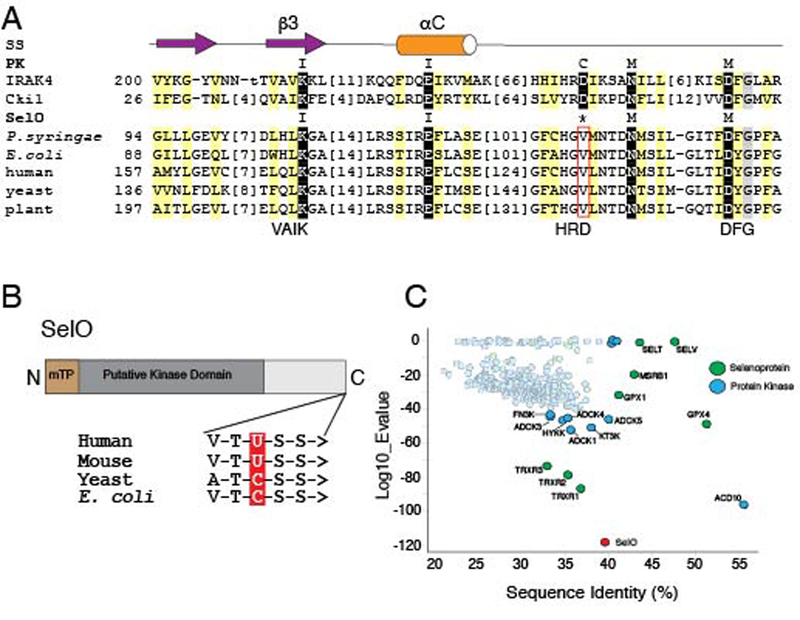
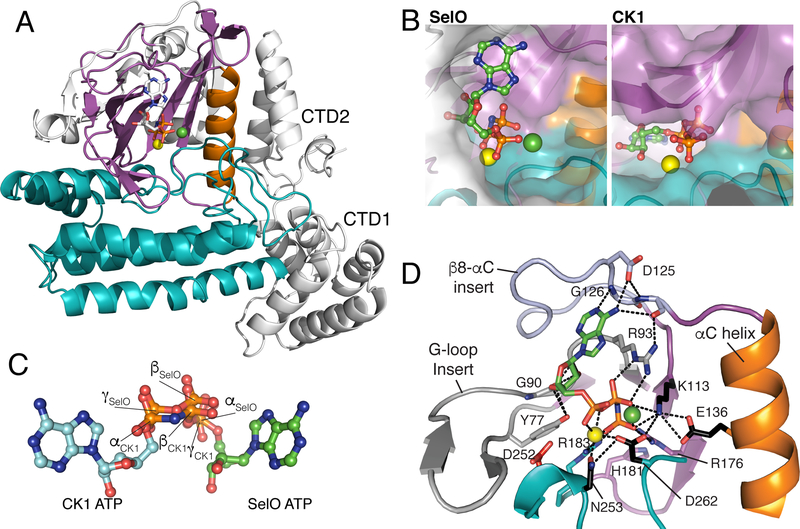
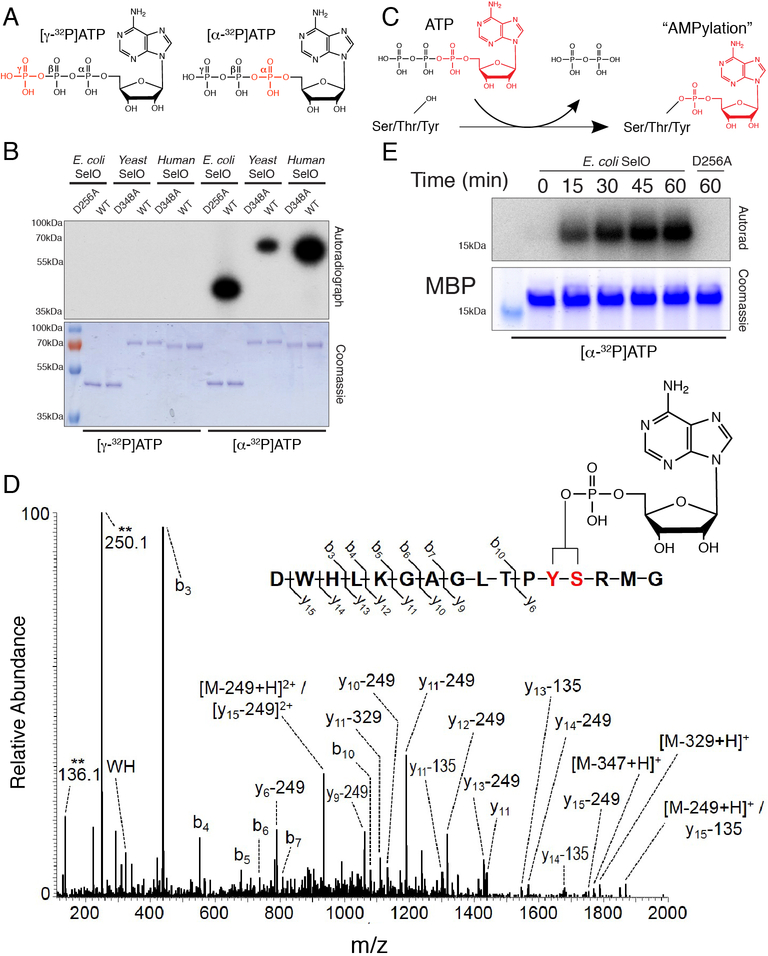
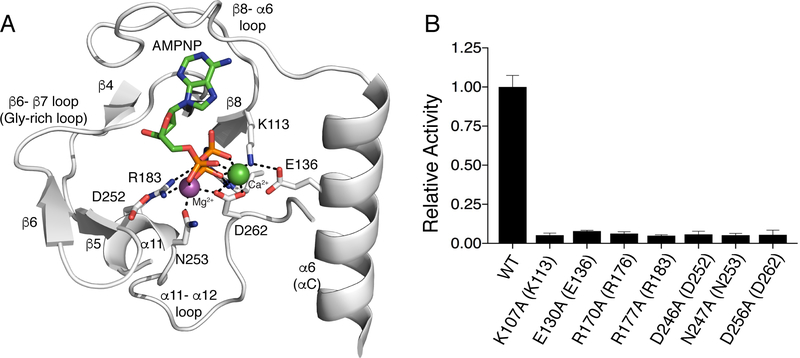
Approximately 10% of human protein kinases are believed to be inactive and named pseudokinases because they lack residues required for catalysis. Here, we show that the highly conserved pseudokinase selenoprotein-O (SelO) transfers AMP from ATP to Ser, Thr, and Tyr residues on protein substrates (AMPylation), uncovering a previously unrecognized activity for a member of the protein kinase superfamily. The crystal structure of a SelO homolog reveals a protein kinase-like fold with ATP flipped in the active site, thus providing a structural basis for catalysis. SelO pseudokinases localize to the mitochondria and AMPylate proteins involved in redox homeostasis. Consequently, SelO activity is necessary for the proper cellular response to oxidative stress. Our results suggest that AMPylation may be a more widespread post-translational modification than previously appreciated and that pseudokinases should be analyzed for alternative transferase activities.
Keywords: SELENOO; adenylylation; glutaredoxin; glutathionylation; kinase structure; oxidative stress; selenocysteine.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures
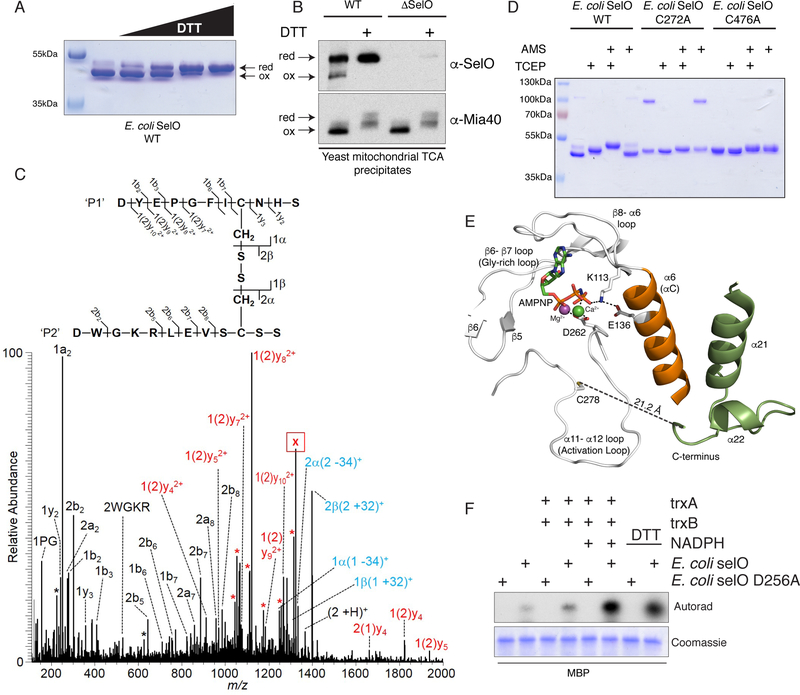
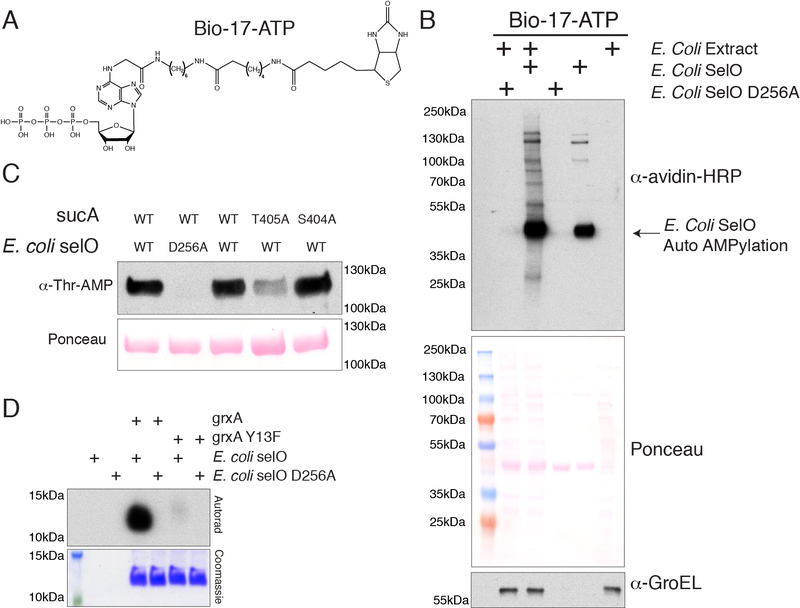
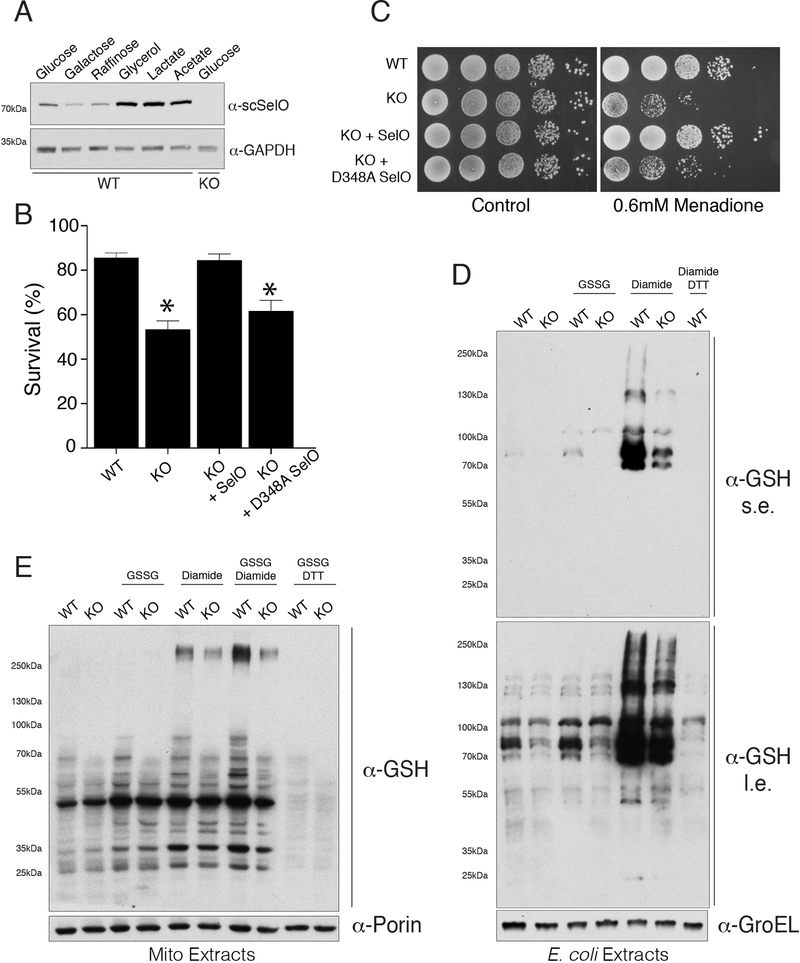
Comment in
-
Flipping ATP to AMPlify Kinase Functions.Cell. 2018 Oct 18;175(3):641-642. doi: 10.1016/j.cell.2018.10.011. Cell. 2018. PMID: 30340038 Free PMC article.
-
Pseudokinases: Flipping the ATP for AMPylation.Curr Biol. 2019 Jan 7;29(1):R23-R25. doi: 10.1016/j.cub.2018.11.018. Curr Biol. 2019. PMID: 30620911
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
