

RIG-I Uses an ATPase-Powered Translocation-Throttling Mechanism for Kinetic Proofreading of RNAs and Oligomerization
- PMID: 30270105
- PMCID: PMC6434538
- DOI: 10.1016/j.molcel.2018.08.021
RIG-I Uses an ATPase-Powered Translocation-Throttling Mechanism for Kinetic Proofreading of RNAs and Oligomerization
Abstract
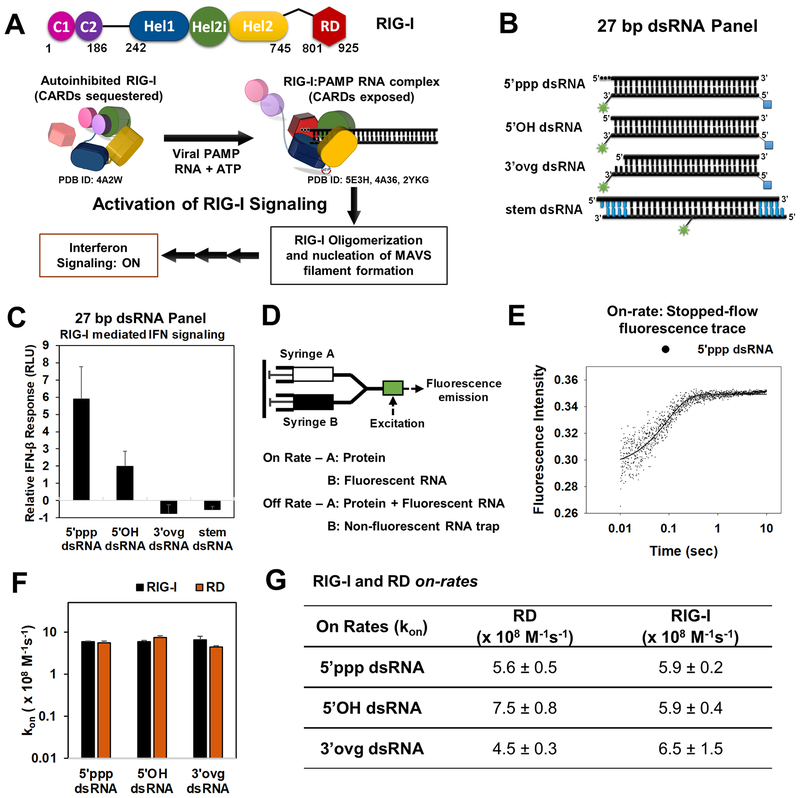
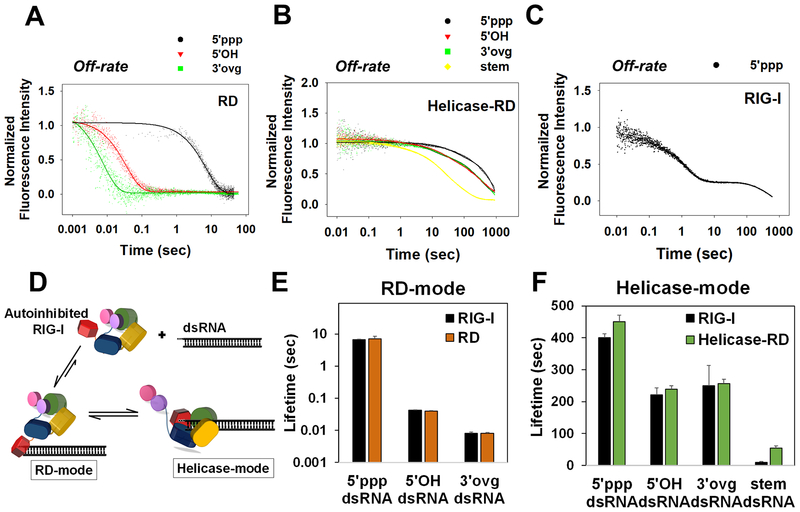
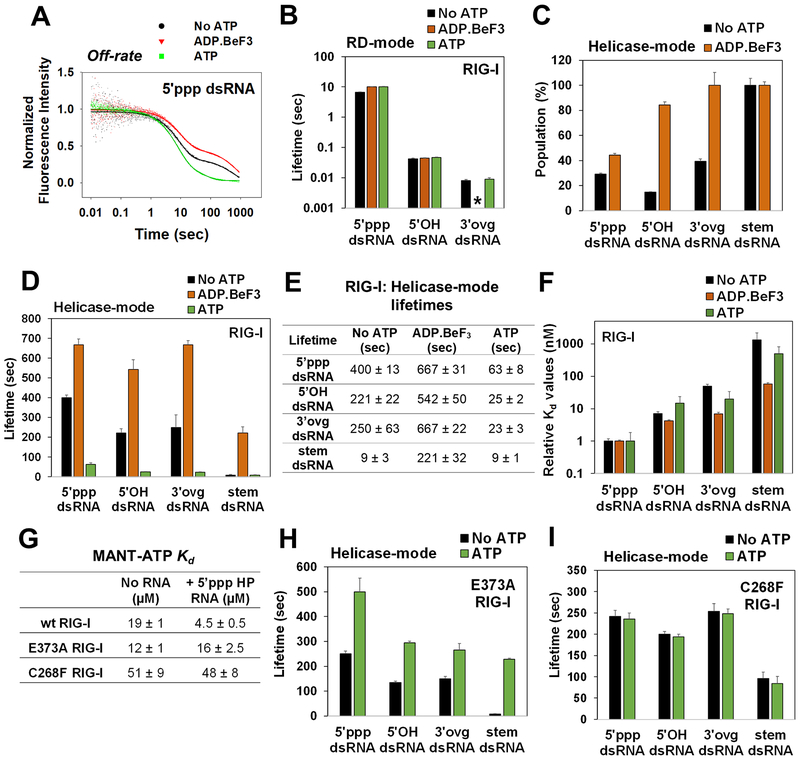
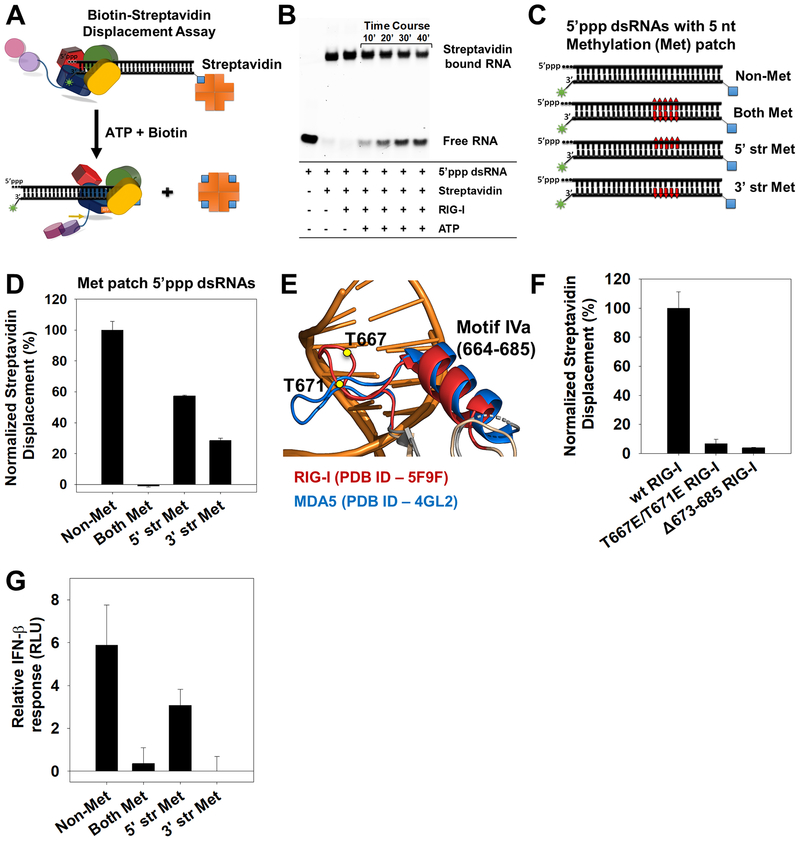
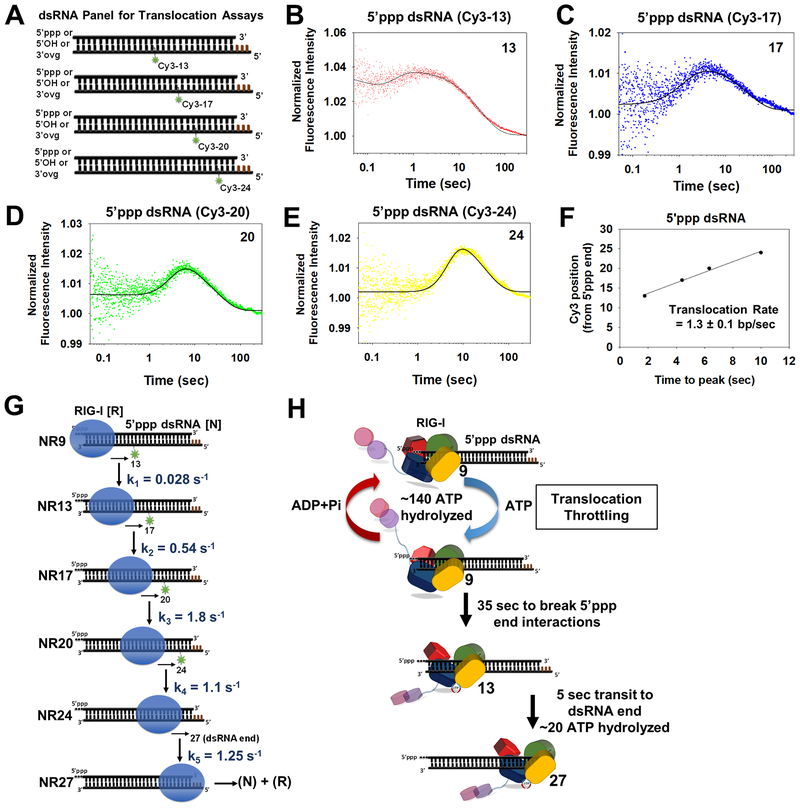
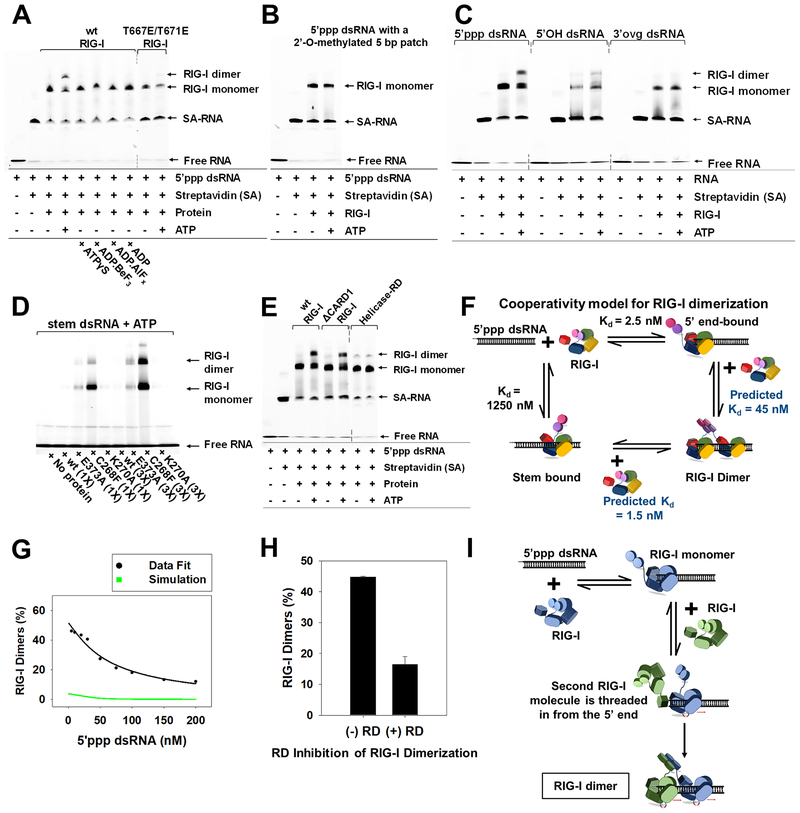
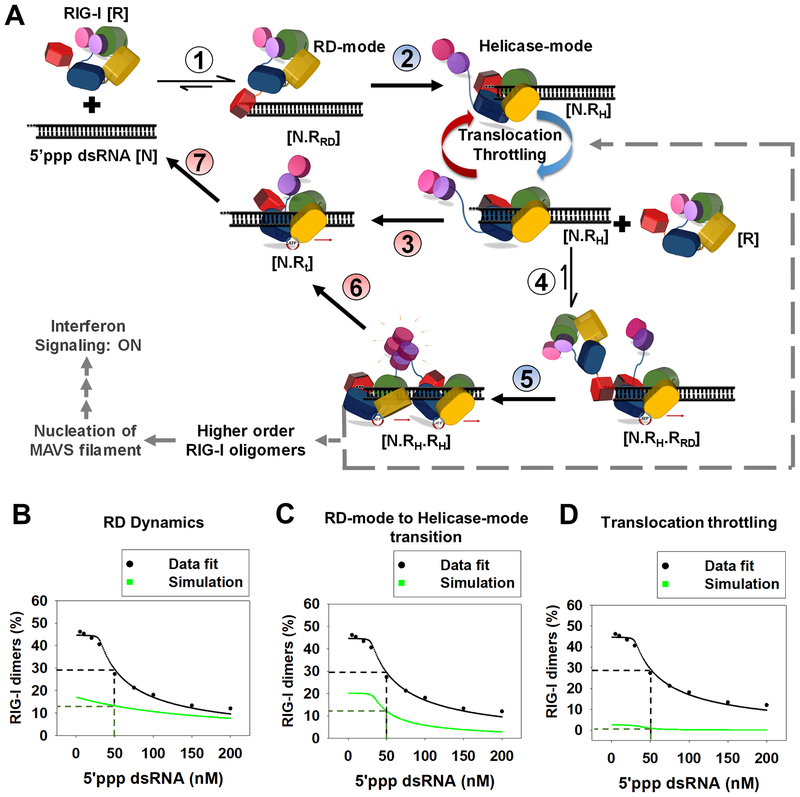
RIG-I has a remarkable ability to specifically select viral 5'ppp dsRNAs for activation from a pool of cytosolic self-RNAs. The ATPase activity of RIG-I plays a role in RNA discrimination and activation, but the underlying mechanism was unclear. Using transient-state kinetics, we elucidated the ATPase-driven "kinetic proofreading" mechanism of RIG-I activation and RNA discrimination, akin to DNA polymerases, ribosomes, and T cell receptors. Even in the autoinhibited state of RIG-I, the C-terminal domain kinetically discriminates against self-RNAs by fast off rates. ATP binding facilitates dsRNA engagement but, interestingly, makes RIG-I promiscuous, explaining the constitutive signaling by Singleton-Merten syndrome-linked mutants that bind ATP without hydrolysis. ATP hydrolysis dissociates self-RNAs faster than 5'ppp dsRNA but, more importantly, drives RIG-I oligomerization through translocation, which we show to be regulated by helicase motif IVa. RIG-I translocates directionally from the dsRNA end into the stem region, and the 5'ppp end "throttles" translocation to provide a mechanism for threading and building a signaling-active oligomeric complex.
Keywords: ATP; RIG-I; RNA discrimination; innate immunity; kinetic proofreading; kinetics; oligomerization; self-versus-non-self; translocation.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures
References
-
- Banerjee K, Kolomeisky AB, and Igoshin OA (2017). Accuracy of Substrate Selection by Enzymes Is Controlled by Kinetic Discrimination. J Phys Chem Lett 8, 1552–1556. - PubMed
-
- Blanchard SC, Gonzalez RL, Kim HD, Chu S, and Puglisi JD (2004). tRNA selection and kinetic proofreading in translation. Nat Struct Mol Biol 11, 1008–1014. - PubMed
-
- Dillingham MS, Wigley DB, and Webb MR (2000). Demonstration of unidirectional single-stranded DNA translocation by PcrA helicase: measurement of step size and translocation speed. Biochemistry 39, 205–212. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
