

Automated Design of Efficient and Functionally Diverse Enzyme Repertoires
- PMID: 30270109
- PMCID: PMC6193528
- DOI: 10.1016/j.molcel.2018.08.033
Automated Design of Efficient and Functionally Diverse Enzyme Repertoires
Abstract
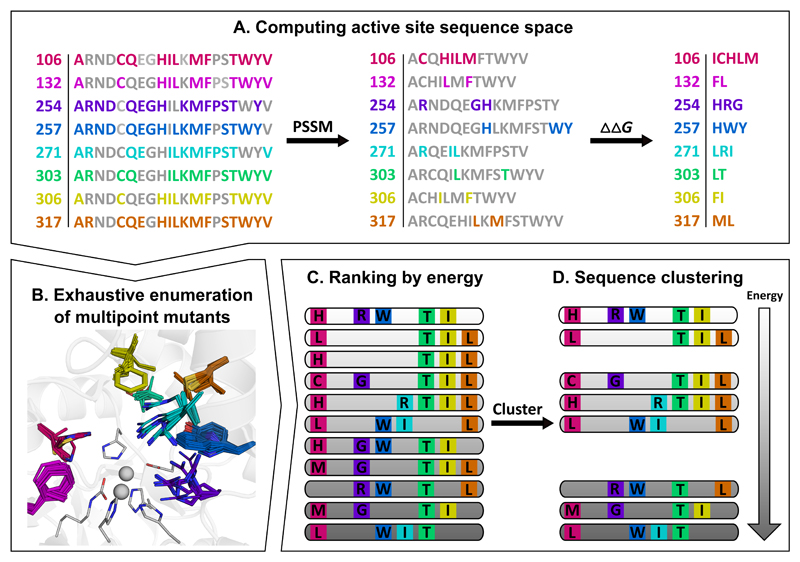
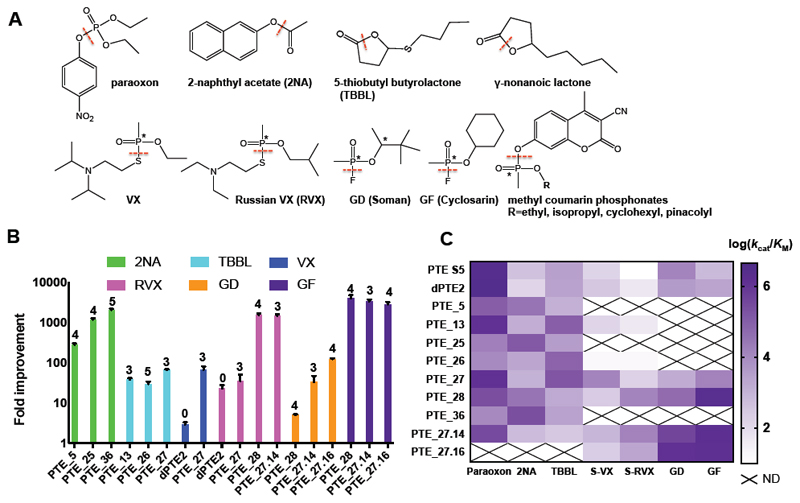
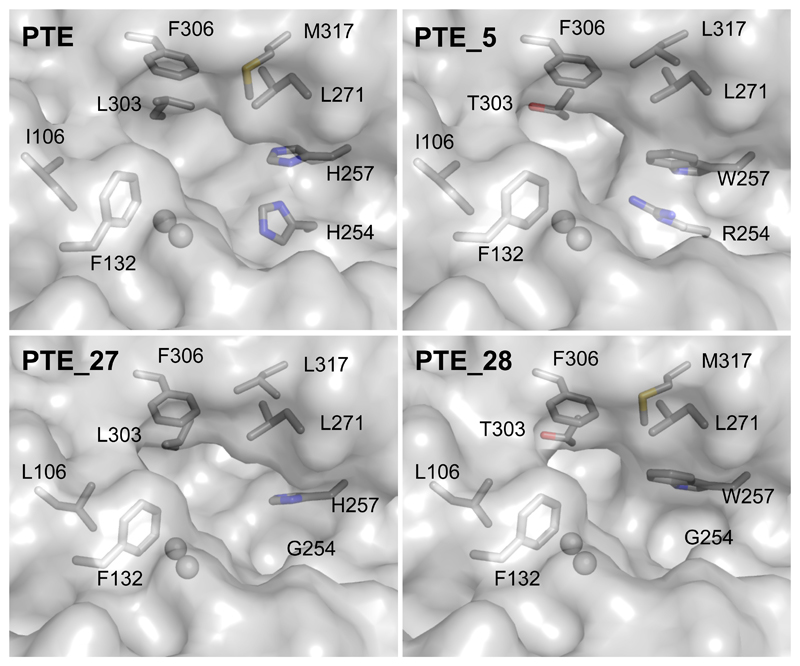
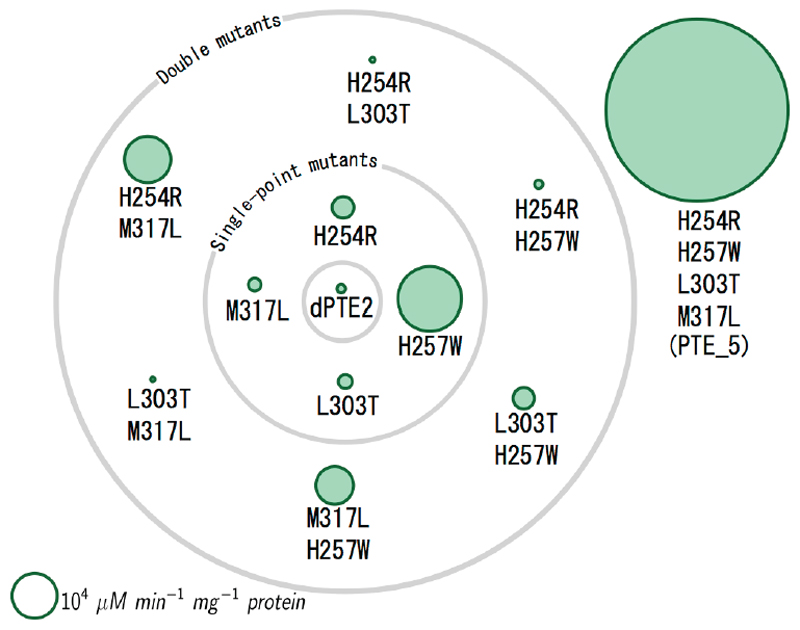
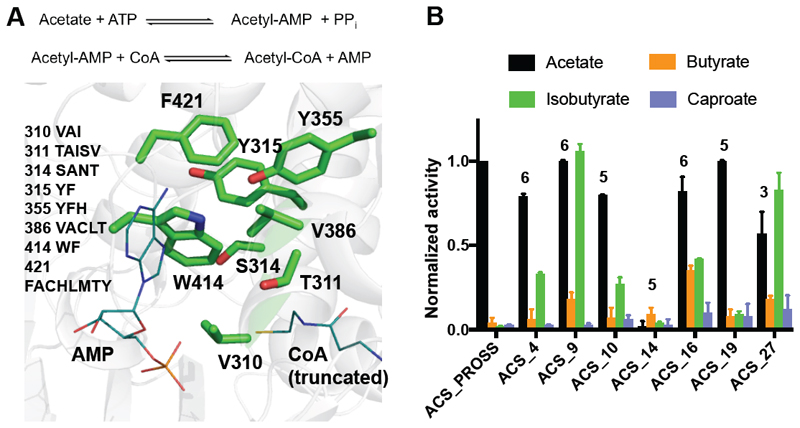
Substantial improvements in enzyme activity demand multiple mutations at spatially proximal positions in the active site. Such mutations, however, often exhibit unpredictable epistatic (non-additive) effects on activity. Here we describe FuncLib, an automated method for designing multipoint mutations at enzyme active sites using phylogenetic analysis and Rosetta design calculations. We applied FuncLib to two unrelated enzymes, a phosphotriesterase and an acetyl-CoA synthetase. All designs were active, and most showed activity profiles that significantly differed from the wild-type and from one another. Several dozen designs with only 3-6 active-site mutations exhibited 10- to 4,000-fold higher efficiencies with a range of alternative substrates, including hydrolysis of the toxic organophosphate nerve agents soman and cyclosarin and synthesis of butyryl-CoA. FuncLib is implemented as a web server (http://FuncLib.weizmann.ac.il); it circumvents iterative, high-throughput experimental screens and opens the way to designing highly efficient and diverse catalytic repertoires.
Keywords: FuncLib; PROSS; Rosetta; enzyme design; enzyme repertoires; epistatic mutations; nerve agents.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Afonine Pavel V, Grosse-Kunstleve Ralf W, Echols Nathaniel, Headd Jeffrey J, Moriarty Nigel W, Mustyakimov Marat, Terwilliger Thomas C, Urzhumtsev Alexandre, Zwart Peter H, Adams Paul D. Towards Automated Crystallographic Structure Refinement with Phenix.refine. Acta Crystallographica. Section D, Biological Crystallography. 2012;68(Pt 4):352–67. - PMC - PubMed
-
- Afriat-Jurnou Livnat, Jackson Colin J, Tawfik Dan S. Reconstructing a Missing Link in the Evolution of a Recently Diverged Phosphotriesterase by Active-Site Loop Remodeling. Biochemistry. 2012;51(31):6047–55. - PubMed
-
- Badenhorst Christoffel PS, Bornscheuer Uwe T. Getting Momentum: From Biocatalysis to Advanced Synthetic Biology. Trends in Biochemical Sciences. 2018;43(3):180–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
