

Coordination between binocular field and spontaneous self-motion specifies the efficiency of planarians' photo-response orientation behavior
- PMID: 30272024
- PMCID: PMC6155068
- DOI: 10.1038/s42003-018-0151-2
Coordination between binocular field and spontaneous self-motion specifies the efficiency of planarians' photo-response orientation behavior
Abstract
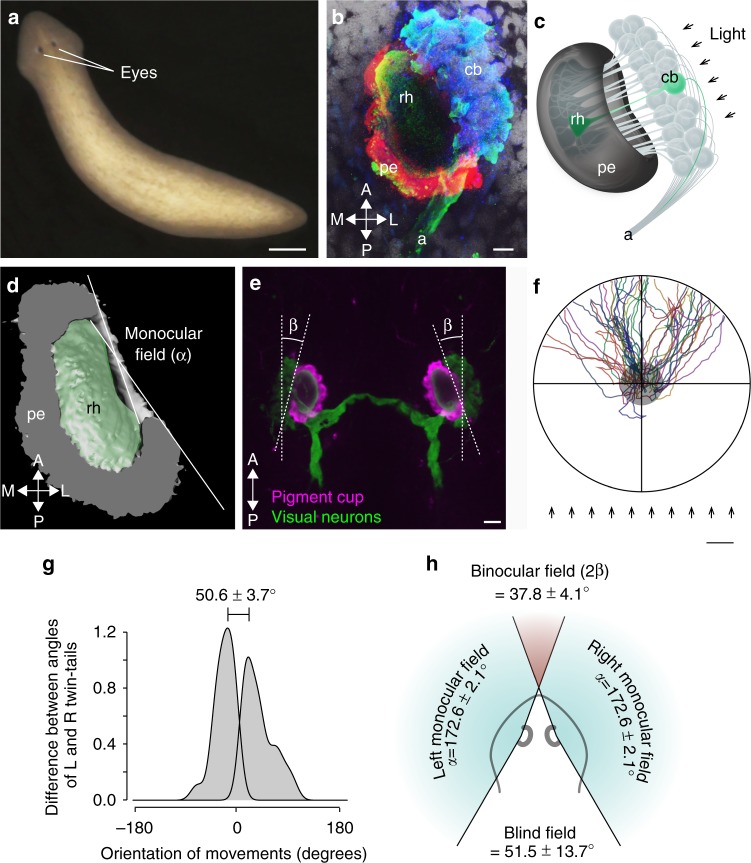
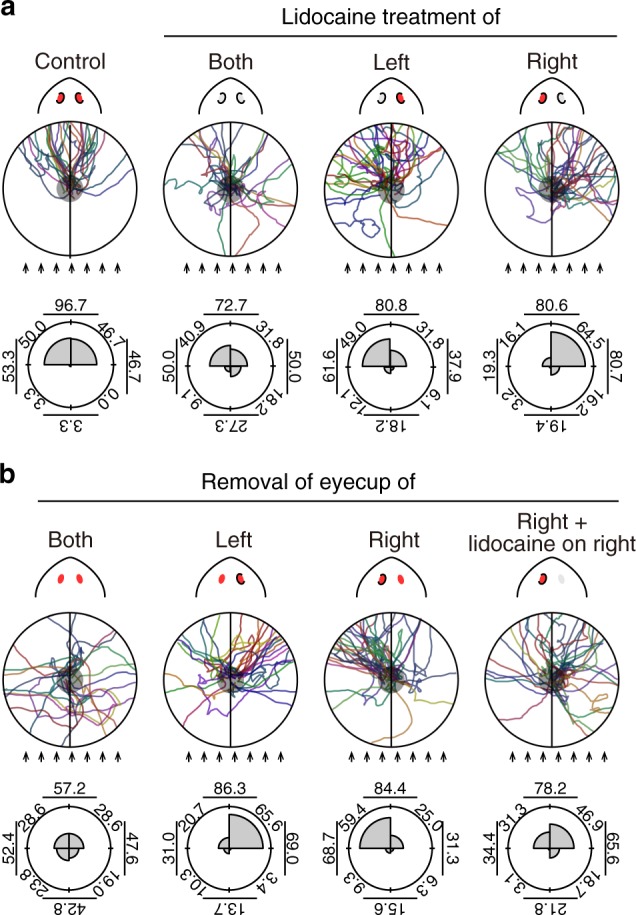
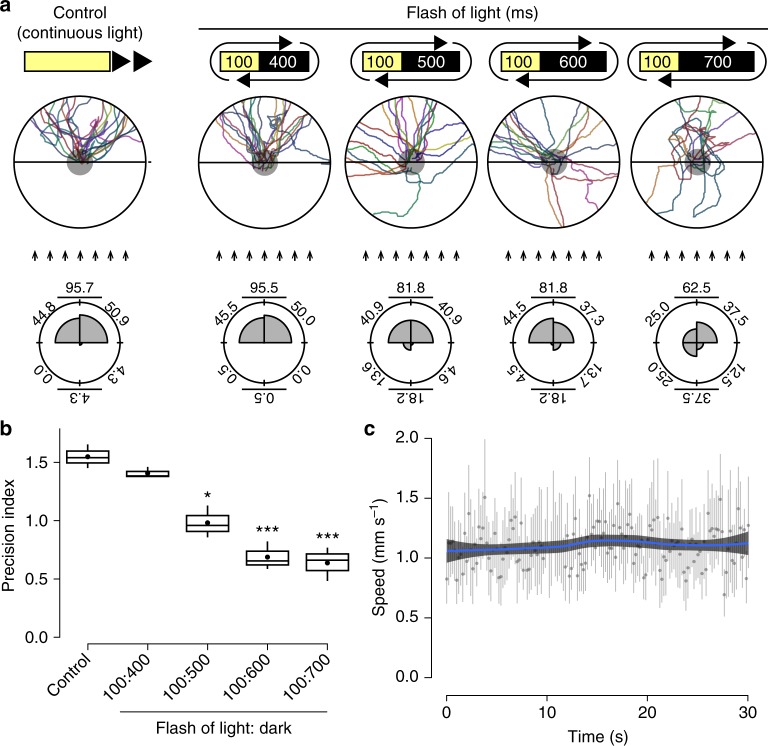
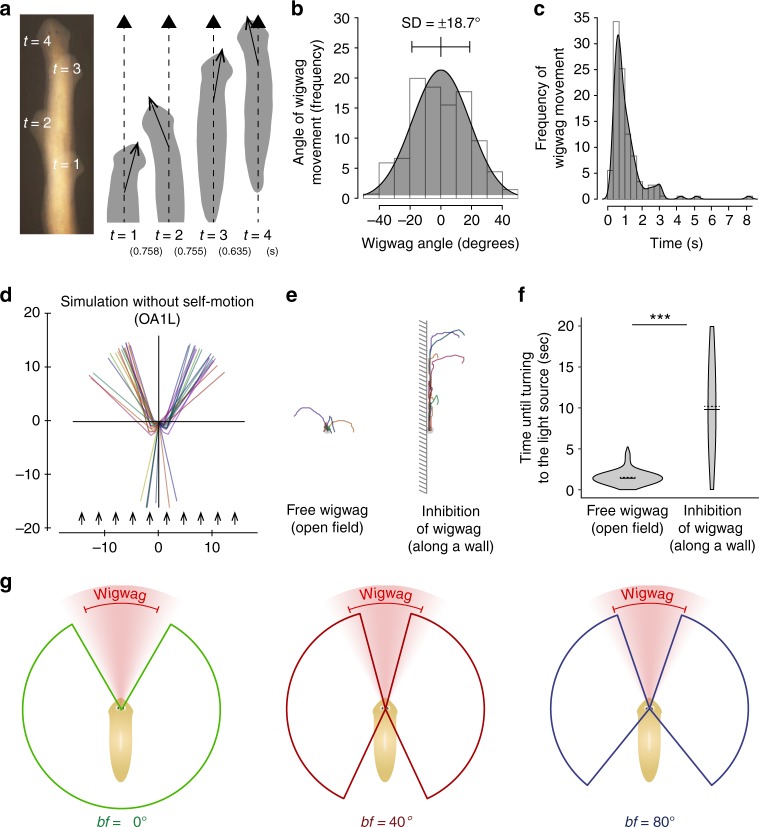
Eyes show remarkable diversity in morphology among creatures. However, little is known about how morphological traits of eyes affect behaviors. Here, we investigate the mechanisms responsible for the establishment of efficient photo-response orientation behavior using the planarian Dugesia japonica as a model. Our behavioral assays reveal the functional angle of the visual field and show that the binocular field formed by paired eyes in D. japonica has an impact on the accurate recognition of the direction of a light source. Furthermore, we find that the binocular field in coordination with spontaneous wigwag self-motion of the head specifies the efficiency of photo-responsive evasive behavior in planarians. Our findings suggest that the linkage between the architecture of the sensory organs and spontaneous self-motion is a platform that serves for efficient and adaptive outcomes of planarian and potentially other animal behaviors.
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Spontaneous Behaviors and Wall-Curvature Lead to Apparent Wall Preference in Planarian.PLoS One. 2015 Nov 5;10(11):e0142214. doi: 10.1371/journal.pone.0142214. eCollection 2015. PLoS One. 2015. PMID: 26539715 Free PMC article.
-
Planarian shows decision-making behavior in response to multiple stimuli by integrative brain function.Zoological Lett. 2015 Feb 1;1:7. doi: 10.1186/s40851-014-0010-z. eCollection 2015. Zoological Lett. 2015. PMID: 26605052 Free PMC article.
-
Pharmacological assessment of methamphetamine-induced behavioral hyperactivity mediated by dopaminergic transmission in planarian Dugesia japonica.Biochem Biophys Res Commun. 2014 Jul 11;449(4):412-8. doi: 10.1016/j.bbrc.2014.05.059. Epub 2014 May 22. Biochem Biophys Res Commun. 2014. PMID: 24858686
-
Regeneration in an evolutionarily primitive brain--the planarian Dugesia japonica model.Eur J Neurosci. 2011 Sep;34(6):863-9. doi: 10.1111/j.1460-9568.2011.07819.x. Eur J Neurosci. 2011. PMID: 21929621 Review.
-
Evolution and regeneration of the planarian central nervous system.Dev Growth Differ. 2009 Apr;51(3):185-95. doi: 10.1111/j.1440-169X.2009.01099.x. Dev Growth Differ. 2009. PMID: 19379275 Review.
Cited by
-
map3k1 suppresses terminal differentiation of migratory eye progenitors in planarian regeneration.PLoS Genet. 2025 Mar 17;21(3):e1011457. doi: 10.1371/journal.pgen.1011457. eCollection 2025 Mar. PLoS Genet. 2025. PMID: 40096024 Free PMC article.
-
Calcium ions in the aquatic environment drive planarians to food.Zoological Lett. 2019 Nov 6;5:31. doi: 10.1186/s40851-019-0147-x. eCollection 2019. Zoological Lett. 2019. PMID: 31720007 Free PMC article.
-
Regeneration of Planarian Auricles and Reestablishment of Chemotactic Ability.Front Cell Dev Biol. 2021 Nov 26;9:777951. doi: 10.3389/fcell.2021.777951. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34901022 Free PMC article.
-
map3k1 suppresses terminal differentiation of migratory eye progenitors in planarian regeneration.bioRxiv [Preprint]. 2024 Oct 12:2024.10.11.617745. doi: 10.1101/2024.10.11.617745. bioRxiv. 2024. Update in: PLoS Genet. 2025 Mar 17;21(3):e1011457. doi: 10.1371/journal.pgen.1011457. PMID: 39416008 Free PMC article. Updated. Preprint.
-
Vasopressin-oxytocin-type signaling is ancient and has a conserved water homeostasis role in euryhaline marine planarians.Sci Adv. 2022 Mar 4;8(9):eabk0331. doi: 10.1126/sciadv.abk0331. Epub 2022 Mar 4. Sci Adv. 2022. PMID: 35245108 Free PMC article.
References
-
- Land Michael F., Nilsson Dan-Eric. Animal Eyes. 2012. Lens eyes on land; pp. 94–129.
-
- Hays GC. A review of the adaptive significance and ecosystem consequences of zooplankton diel vertical migrations. Hydrobiologia. 2003;503:163–170. doi: 10.1023/B:HYDR.0000008476.23617.b0. - DOI
-
- Togashi T, Cox PA. Phototaxis and the evolution of isogamy and “slight anisogamy” in marine green algae: insights from laboratory observations and numerical experiments. Bot. J. Linn. Soc. 2004;144:321–327. doi: 10.1111/j.1095-8339.2003.00255.x. - DOI
LinkOut - more resources
Full Text Sources
