

Fenofibrate Interferes with the Diapedesis of Lung Adenocarcinoma Cells through the Interference with Cx43/EGF-Dependent Intercellular Signaling
- PMID: 30274176
- PMCID: PMC6210471
- DOI: 10.3390/cancers10100363
Fenofibrate Interferes with the Diapedesis of Lung Adenocarcinoma Cells through the Interference with Cx43/EGF-Dependent Intercellular Signaling
Abstract
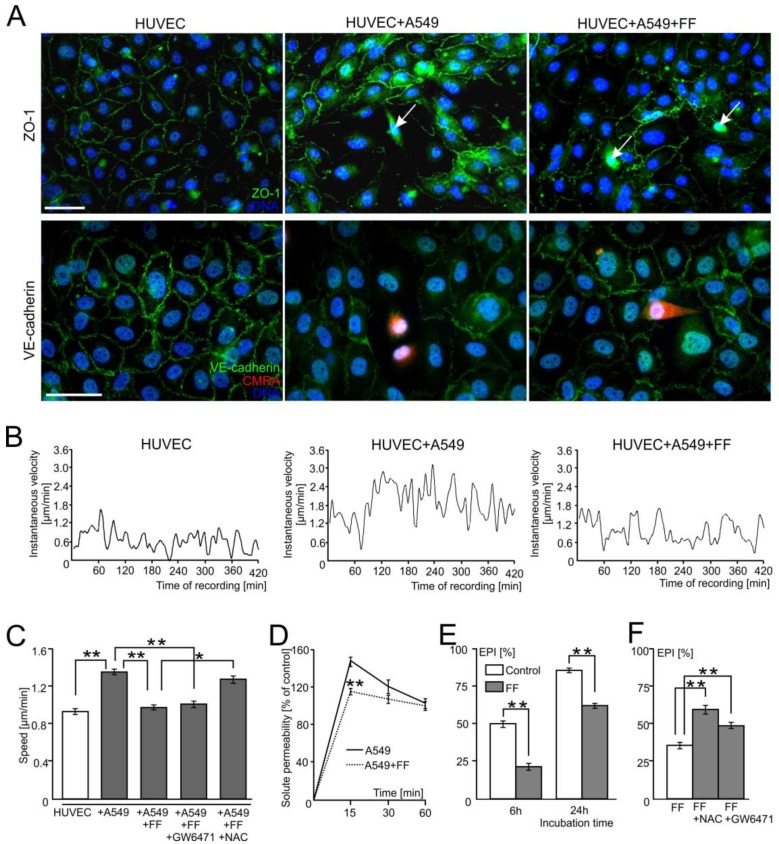
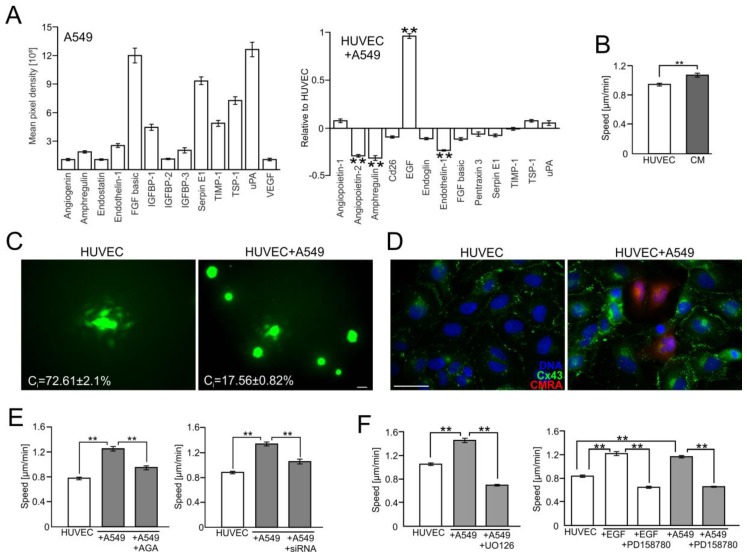
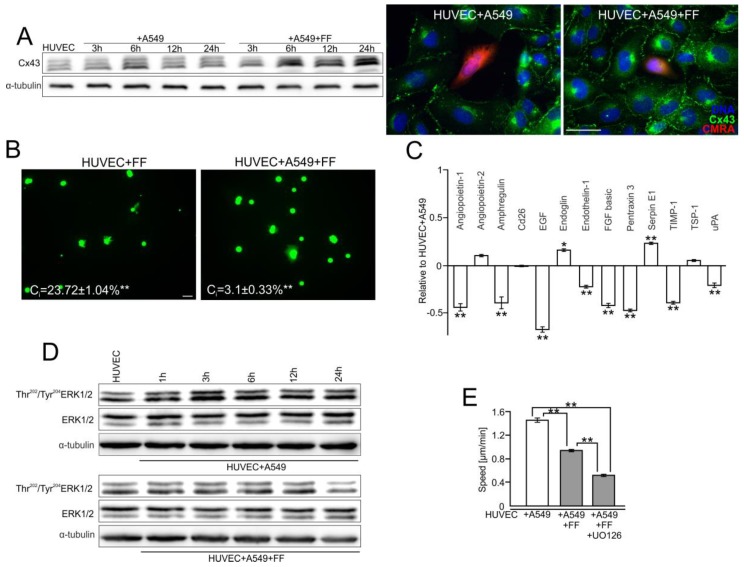
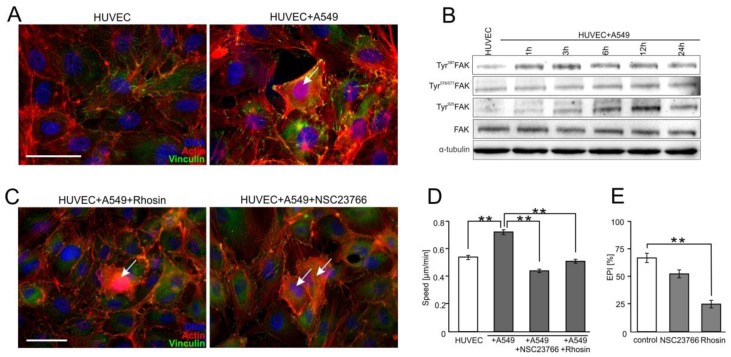
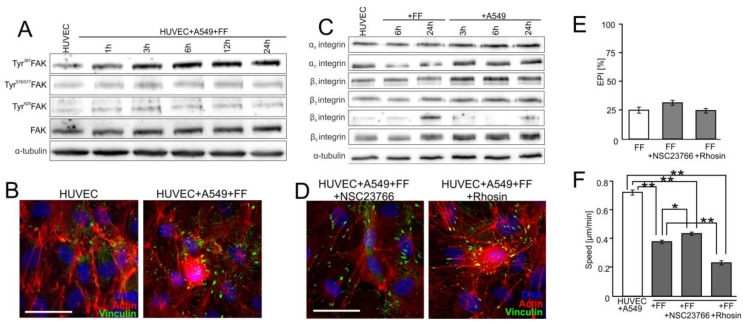
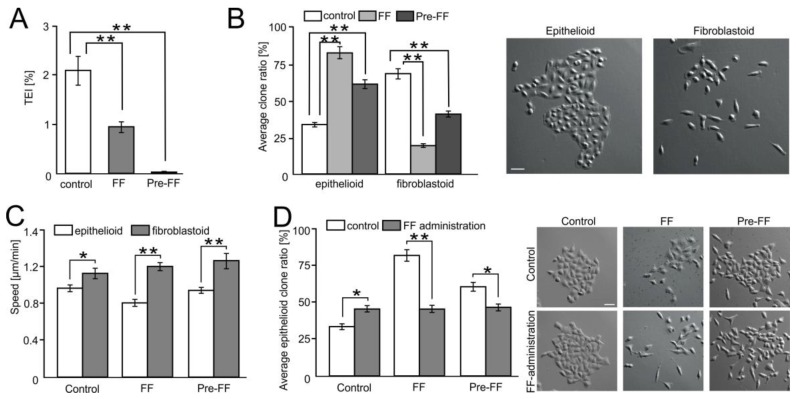
Extravasation of circulating cancer cells is regulated by the intercellular/intracellular signaling pathways that locally impair the endothelial barrier function. Co-cultures of human umbilical vein endothelial cells (HUVECs) with lung adenocarcinoma A549 cells enabled us to identify these pathways and to quantify the effect of fenofibrate (FF) on their activity. A549 cells induced the disruption and local activation of endothelial continuum. These events were accompanied by epidermal growth factor (EGF) up-regulation in endothelial cells. Impaired A549 diapedesis and HUVEC activation were seen upon the chemical inhibition of connexin(Cx)43 functions, EGF/ERK1/2-dependent signaling, and RhoA/Rac1 activity. A total of 25 μM FF exerted corresponding effects on Cx43-mediated gap junctional coupling, EGF production, and ERK1/2 activation in HUVEC/A549 co-cultures. It also directly augmented endothelial barrier function via the interference with focal adhesion kinase (FAK)/RhoA/Rac1-regulated endothelial cell adhesion/contractility/motility and prompted the selective transmigration of epithelioid A549 cells. N-acetyl-L-cysteine abrogated FF effects on HUVEC activation, suggesting the involvement of PPARα-independent mechanism(s) in its action. Our data identify a novel Cx43/EGF/ERK1/2/FAK/RhoA/Rac1-dependent signaling axis, which determines the efficiency of lung cancer cell diapedesis. FF interferes with its activity and reduces the susceptibility of endothelial cells to A549 stimuli. These findings provide the rationale for the implementation of FF in the therapy of malignant lung cancers.
Keywords: Cx43; EGF; diapedesis; fenofibrate; lung cancer.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
