

TET2 Deficiency Causes Germinal Center Hyperplasia, Impairs Plasma Cell Differentiation, and Promotes B-cell Lymphomagenesis
- PMID: 30274972
- PMCID: PMC6279514
- DOI: 10.1158/2159-8290.CD-18-0657
TET2 Deficiency Causes Germinal Center Hyperplasia, Impairs Plasma Cell Differentiation, and Promotes B-cell Lymphomagenesis
Abstract
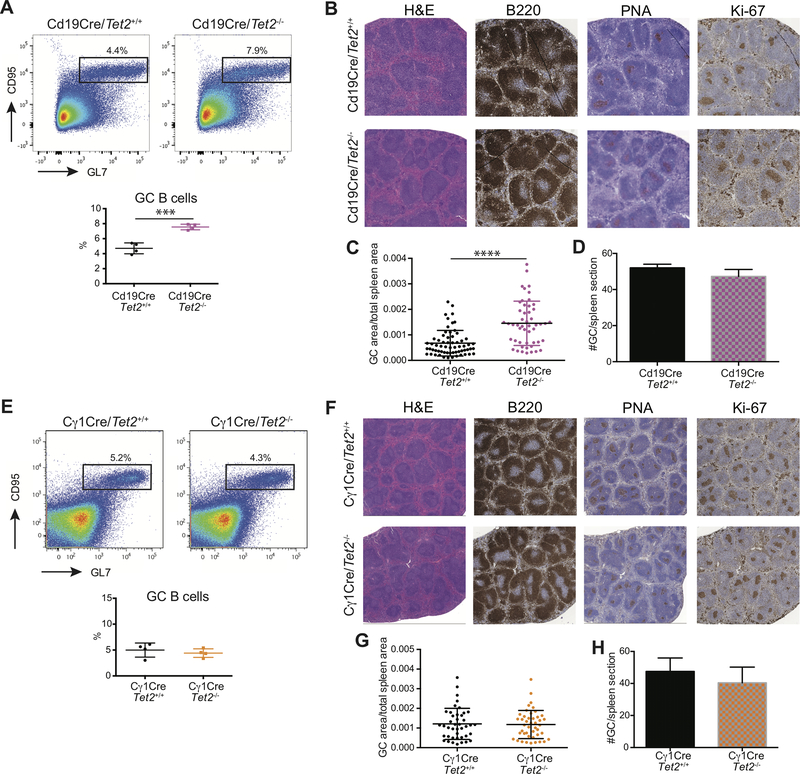
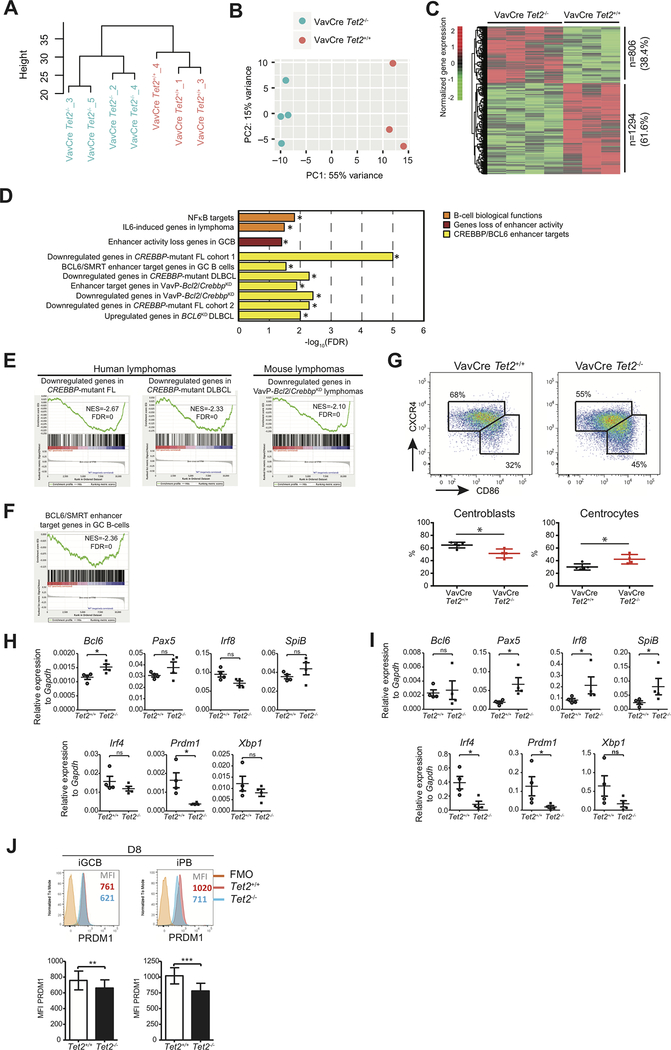
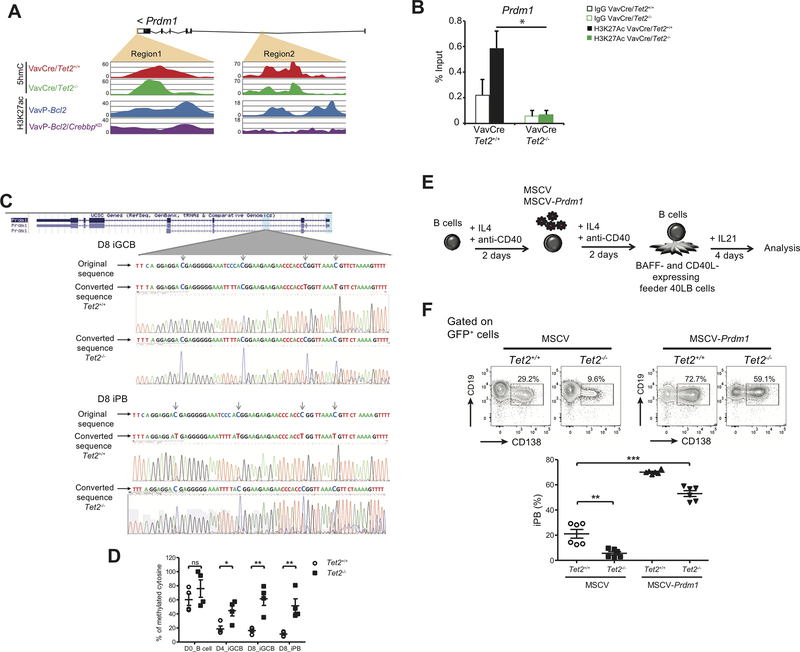
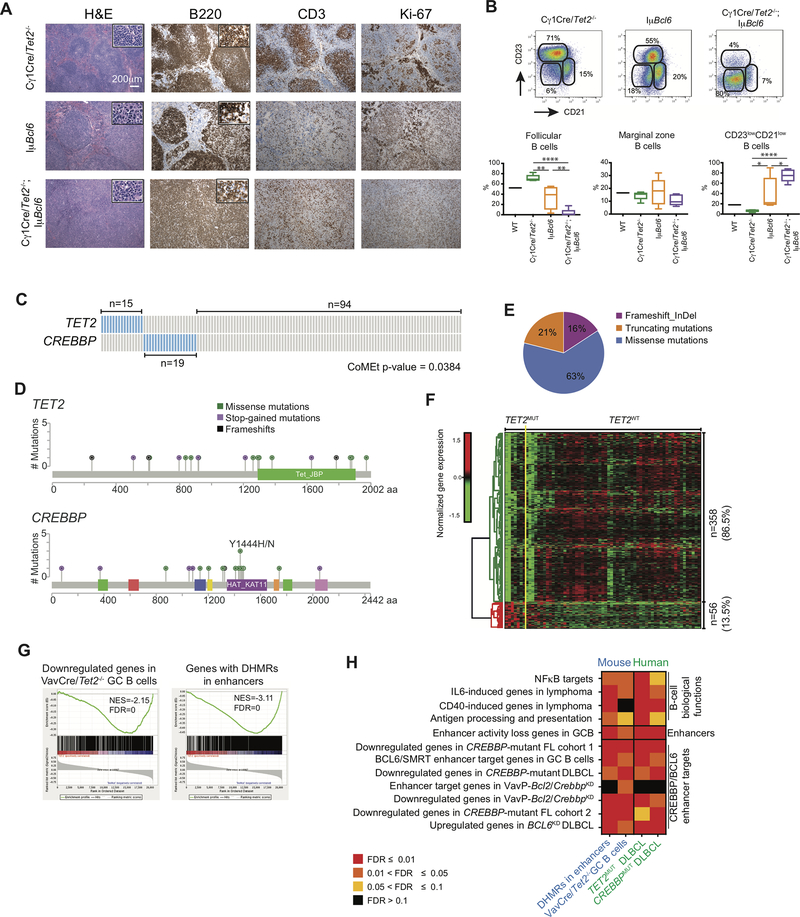
TET2 somatic mutations occur in ∼10% of diffuse large B-cell lymphomas (DLBCL) but are of unknown significance. Herein, we show that TET2 is required for the humoral immune response and is a DLBCL tumor suppressor. TET2 loss of function disrupts transit of B cells through germinal centers (GC), causing GC hyperplasia, impaired class switch recombination, blockade of plasma cell differentiation, and a preneoplastic phenotype. TET2 loss was linked to focal loss of enhancer hydroxymethylation and transcriptional repression of genes that mediate GC exit, such as PRDM1. Notably, these enhancers and genes are also repressed in CREBBP-mutant DLBCLs. Accordingly, TET2 mutation in patients yields a CREBBP-mutant gene-expression signature, CREBBP and TET2 mutations are generally mutually exclusive, and hydroxymethylation loss caused by TET2 deficiency impairs enhancer H3K27 acetylation. Hence, TET2 plays a critical role in the GC reaction, and its loss of function results in lymphomagenesis through failure to activate genes linked to GC exit signals. SIGNIFICANCE: We show that TET2 is required for exit of the GC, B-cell differentiation, and is a tumor suppressor for mature B cells. Loss of TET2 phenocopies CREBBP somatic mutation. These results advocate for sequencing TET2 in patients with lymphoma and for the testing of epigenetic therapies to treat these tumors.See related commentary by Shingleton and Dave, p. 1515.This article is highlighted in the In This Issue feature, p. 1494.
©2018 American Association for Cancer Research.
Conflict of interest statement
Disclosure of Potential Conflict of Interest
A. Melnick reports receiving research support from Janssen. R. Levine is in the supervisory board of Qiagen, SAB for Loxo, C4 therapeutics and Isoplexis; and he is a consultant and receives research support from Roche and Celgene. No potential conflicts of interest were disclosed by the other authors.
Figures



Comment in
-
TET2 Deficiency Sets the Stage for B-cell Lymphoma.Cancer Discov. 2018 Dec;8(12):1515-1517. doi: 10.1158/2159-8290.CD-18-1143. Cancer Discov. 2018. PMID: 30510015
References
-
- Coiffier B, Thieblemont C, Van Den Neste E, Lepeu G, Plantier I, Castaigne S, et al. Long-term outcome of patients in the LNH-98.5 trial, the first randomized study comparing rituximab-CHOP to standard CHOP chemotherapy in DLBCL patients: a study by the Groupe d’Etudes des Lymphomes de l’Adulte. Blood 2010;116(12):2040–5 doi . - DOI - PMC - PubMed
-
- Klose RJ, Bird AP. Genomic DNA methylation: the mark and its mediators. Trends Biochem Sci 2006. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
