

The Story of RNA Folding, as Told in Epochs
- PMID: 30275276
- PMCID: PMC6169817
- DOI: 10.1101/cshperspect.a032433
The Story of RNA Folding, as Told in Epochs
Abstract
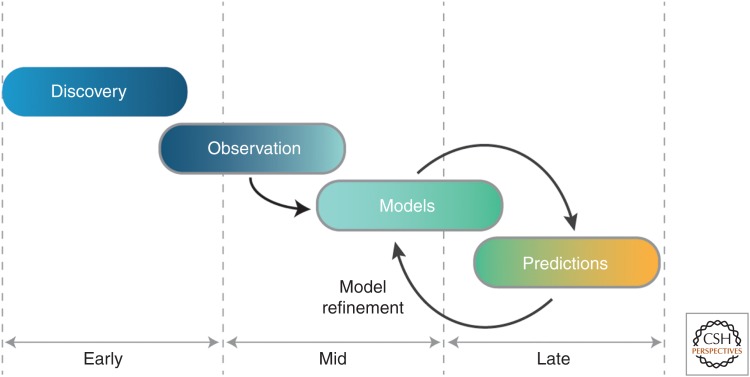
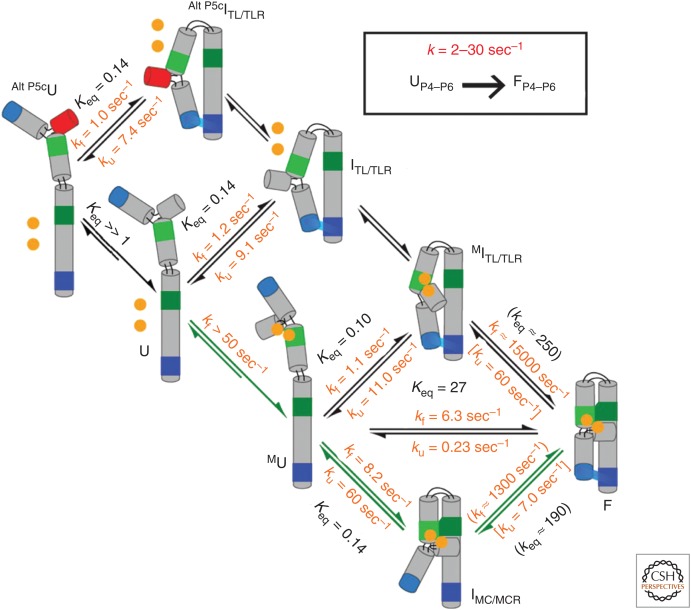
The past decades have witnessed tremendous developments in our understanding of RNA biology. At the core of these advances have been studies aimed at discerning RNA structure and at understanding the forces that influence the RNA folding process. It is easy to take the present state of understanding for granted, but there is much to be learned by considering the path to our current understanding, which has been tortuous, with the birth and death of models, the adaptation of experimental tools originally developed for characterization of protein structure and catalysis, and the development of novel tools for probing RNA. In this review we tour the stages of RNA folding studies, considering them as "epochs" that can be generalized across scientific disciplines. These epochs span from the discovery of catalytic RNA, through biophysical insights into the putative primordial RNA World, to characterization of structured RNAs, the building and testing of models, and, finally, to the development of models with the potential to yield generalizable predictive and quantitative models for RNA conformational, thermodynamic, and kinetic behavior. We hope that this accounting will aid others as they navigate the many fascinating questions about RNA and its roles in biology, in the past, present, and future.
Copyright © 2018 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures






References
-
- Bailor MH, Sun X, Al-Hashimi HM. 2010. Topology links RNA secondary structure with global conformation, dynamics, and adaptation. Science 327: 202–206. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous