

Coassembly of SecYEG and SecA Fully Restores the Properties of the Native Translocon
- PMID: 30275279
- PMCID: PMC6287467
- DOI: 10.1128/JB.00493-18
Coassembly of SecYEG and SecA Fully Restores the Properties of the Native Translocon
Abstract
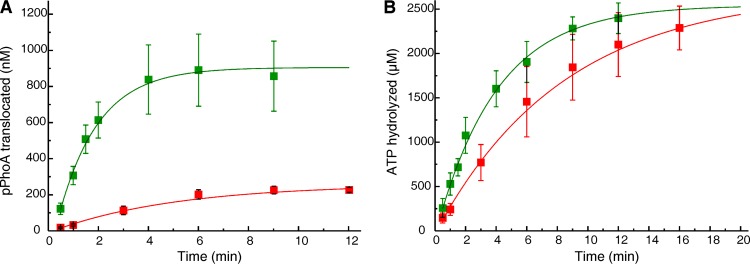
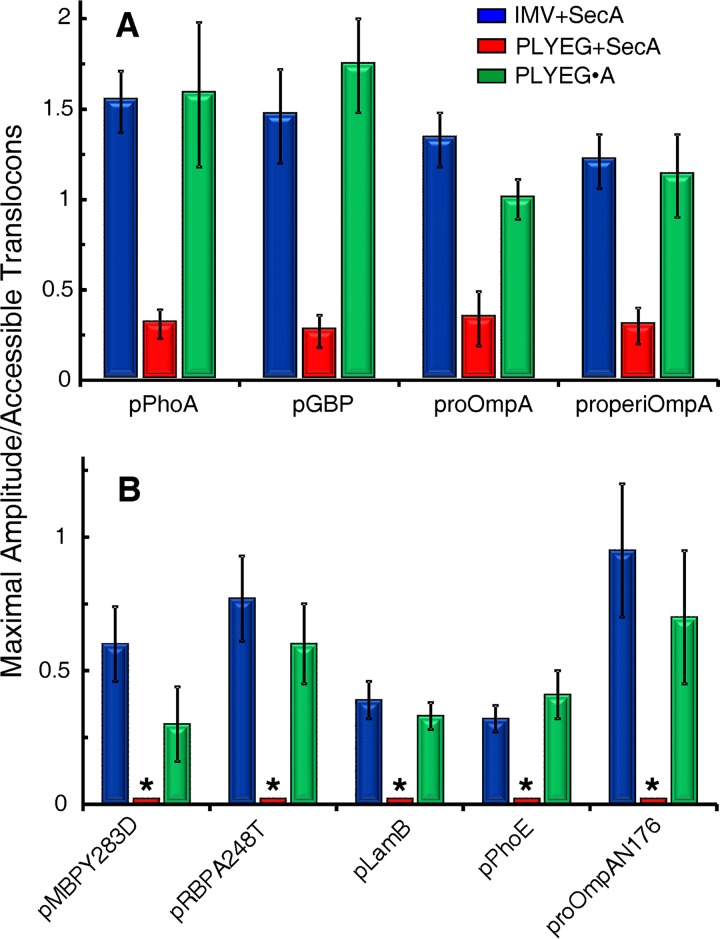
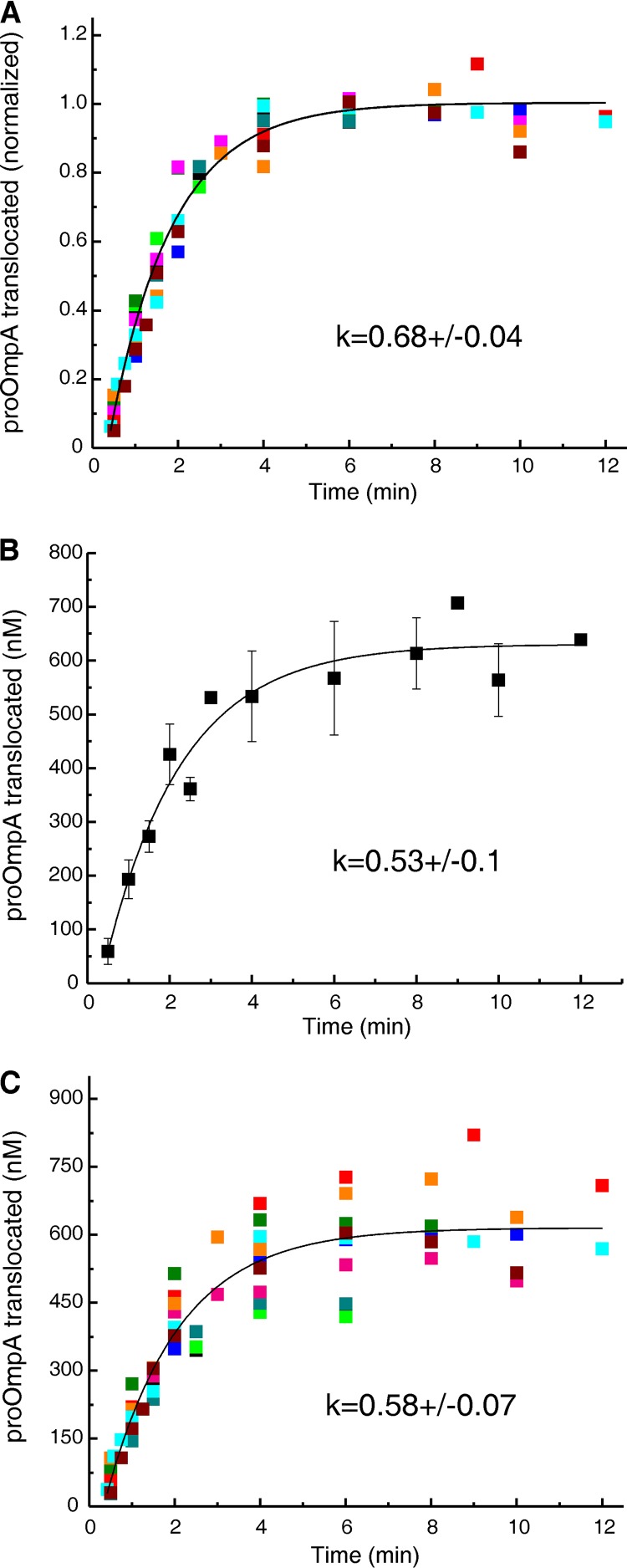
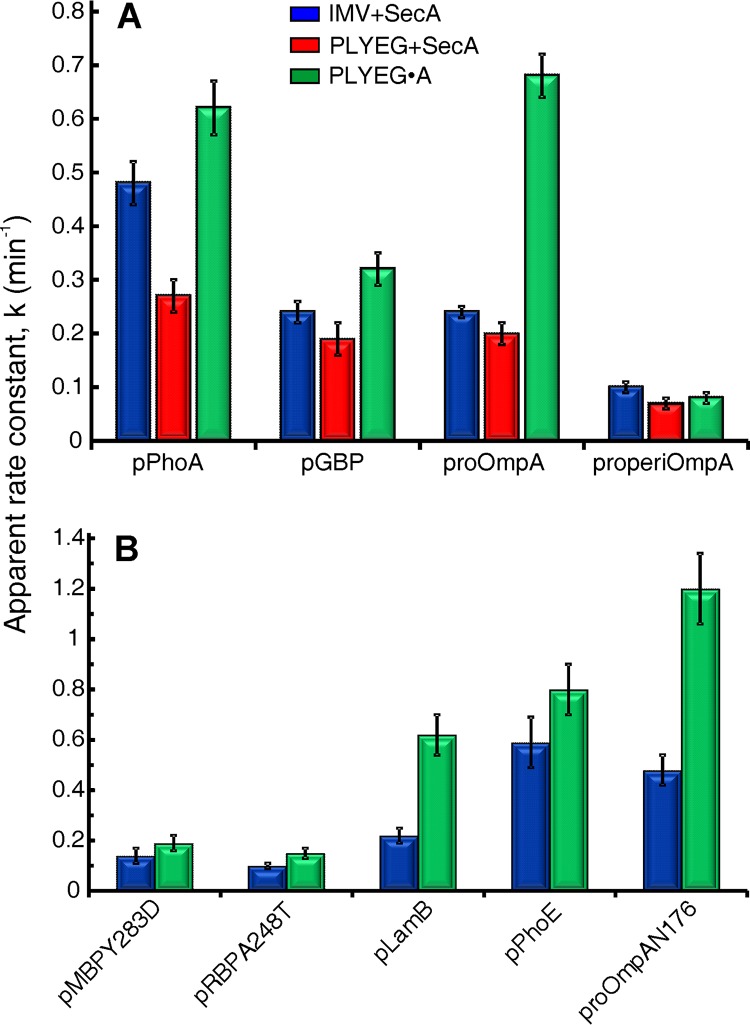
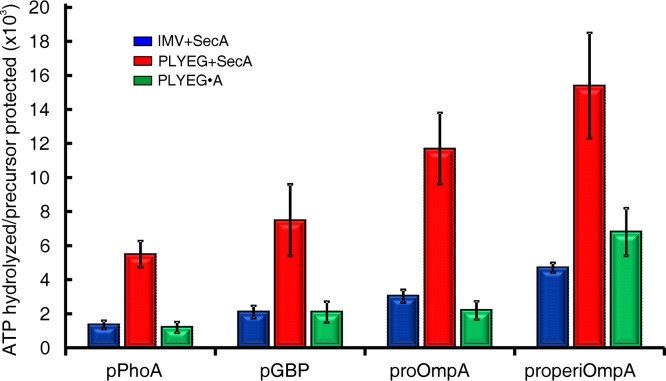
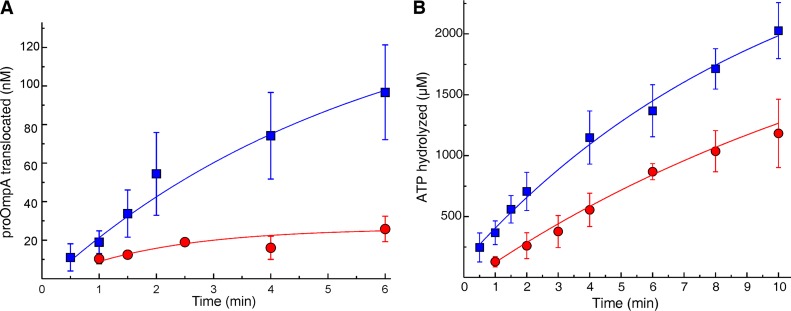
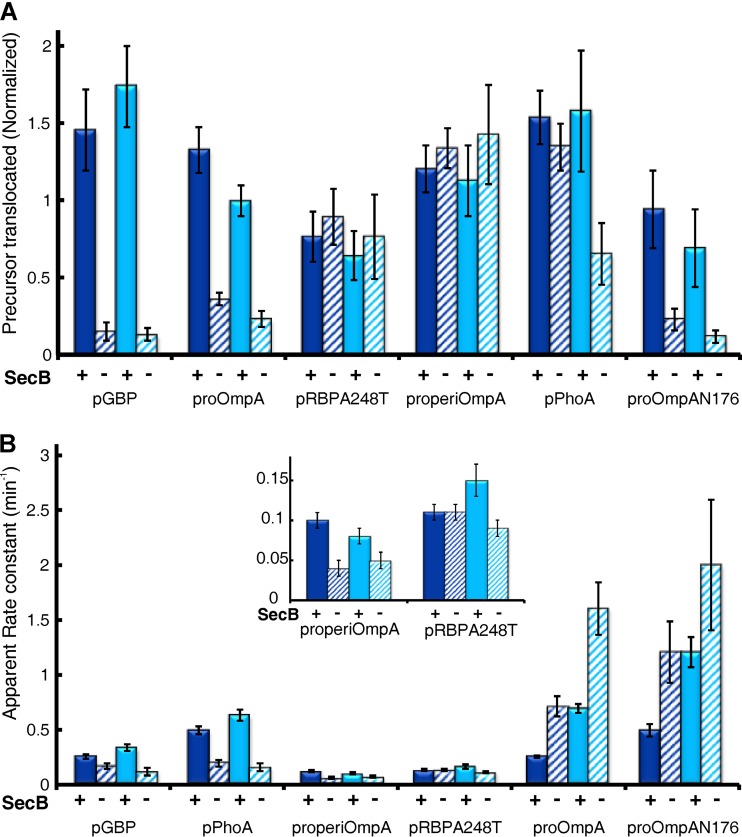
In all cells, a highly conserved channel transports proteins across membranes. In Escherichia coli, that channel is SecYEG. Many investigations of this protein complex have used purified SecYEG reconstituted into proteoliposomes. How faithfully do activities of reconstituted systems reflect the properties of SecYEG in the native membrane environment? We investigated by comparing three in vitro systems: the native membrane environment of inner membrane vesicles and two methods of reconstitution. One method was the widely used reconstitution of SecYEG alone into lipid bilayers. The other was our method of coassembly of SecYEG with SecA, the ATPase of the translocase. For nine different precursor species we assessed parameters that characterize translocation: maximal amplitude of competent precursor translocated, coupling of energy to transfer, and apparent rate constant. In addition, we investigated translocation in the presence and absence of chaperone SecB. For all nine precursors, SecYEG coassembled with SecA was as active as SecYEG in native membrane for each of the parameters studied. Effects of SecB on transport of precursors faithfully mimicked observations made in vivo From investigation of the nine different precursors, we conclude that the apparent rate constant, which reflects the step that limits the rate of translocation, is dependent on interactions with the translocon of portions of the precursors other than the leader. In addition, in some cases the rate-limiting step is altered by the presence of SecB. Candidates for the rate-limiting step that are consistent with our data are discussed.IMPORTANCE This work presents a comprehensive quantification of the parameters of transport by the Sec general secretory system in the three in vitro systems. The standard reconstitution used by most investigators can be enhanced to yield six times as many active translocons simply by adding SecA to SecYEG during reconstitution. This robust system faithfully reflects the properties of translocation in native membrane vesicles. We have expanded the number of precursors studied to nine. This has allowed us to conclude that the rate constant for translocation varies with precursor species.
Keywords: E. coli; SecA; SecYEG; membrane protein; protein export; protein translocation; proteoliposomes; secretion; translocon.
Copyright © 2018 American Society for Microbiology.
Figures
Comment in
-
Substrate Proteins Take Shape at an Improved Bacterial Translocon.J Bacteriol. 2018 Dec 7;201(1):e00618-18. doi: 10.1128/JB.00618-18. Print 2019 Jan 1. J Bacteriol. 2018. PMID: 30322856 Free PMC article.
References
-
- Cabelli RJ, Dolan KM, Qian LP, Oliver DB. 1991. Characterization of membrane-associated and soluble states of SecA protein from wild-type and SecA51(TS) mutant strains of Escherichia coli. J Biol Chem 266:24420–24427. - PubMed
-
- Matsuyama S-I, Mizushima S. 1995. Biochemical analyses of components comprising the protein translocation machinery of Escherichia coli. Adv Cell Mol Biol Membr Organelles 4:61–84. doi:10.1016/S1874-5172(06)80007-6. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
